博文
能流进化论——生命起源
||
在探讨生命是如何起源这一问题之前,我们先从能流的角度探讨一下汽车的起源问题。
已知A地与B地之间的距离是1900公里,且A地有个人因为一件事而不得不去B地。如果选择步行,按时速5公里,每天步行12小时算,那么这个人需要走30天;如果选择骑自行车,按时速10公里,每天骑行12小时算,至少需要骑行15天;如果选择驾驶汽车,按时速100公里,每天开车12小时算,两天就可以到达B地。由此可见,交通工具的选择与事情完成的效率密切相关。
假如事情必须要在5天内完成,且已知此人选择了汽车作为交通工具并顺利完成任务。对于这个事件,我们可以说,因为汽车的存在,此人才得以在5天内完成任务;也可以说,因为事情必须要再5天内完成,所以此人不得不选择能够帮助他在5天内到达B地的运送能力,而汽车恰好具有此能力。对于这两种说法,貌似都合情合理,可是哪一种更准确呢?很显然是第二种,这是因为,如果当时还有高铁、飞机等时速更快的交通工具可以选择,那么,此人有可能会选择其它种类的交通工具。这个事例说明,“事情必须在5天内完成”是交通工具选择的决定因素,此人最终选择“汽车”只是因为汽车是在满足条件的各种交通工具中相对更适合的一种而已。
既然存在“汽车”可供选择,说明汽车已经存在了。现在退一步讲,汽车又是怎么来的呢?从能流的角度分析,汽车的出现需要解决以下三个问题:
1)要存在可以转化为动能的能流。已知发动机可以将汽油中的化学能转换成动能,意味着利用发动机提供动能的汽车出现之前,首先应该存在发动机。从能流的角度讲,汽车出现之前,首先需要存在“化学能→动能”这样一种能流,汽车的产生是建立在这种能流之上的。从能流的构成分析,汽油是这条能流的能量供体,发动机是这条能流能量传递介体,车轮是这条能流能量受体,因而介导这条能流的能量传递单元记作为“汽油→发动机→车轮”;
2)动能的维持要得到保障。如果需要汽车的动能持续性产生,那么“汽油→发动机→车轮”这条能量传递单元就需要持续性工作。在能量的流动过程中,发动机和车轮可以一直稳定的存在,而汽油却在不断的被消耗。因此,要想保障汽车动能的持续性,就需要保障汽油的量,具体措施包括将发动机与一个油箱相连接,以及修建加油站等。此外,单纯的将汽油、发动机和车轮相连接并不能将汽油中的化学能转换为动能,这是因为该能流的运行还需要活塞的动能、点火系统的热能等其它能量的参与,而这些能量又来源于其它能流,说明“汽油→发动机→车轮”这条能量传递单元的工作还需要其它能量传递单元或能流的辅助。
3)“汽油→发动机→车轮”这条能量传递单元的能量传递效率要足够高。在能量的流动过程中,汽油中的化学能并不会完全转换成车轮的动能,有些还会以热能的形式流入环境中,也有的会转化成其它形式的能量,例如,转化为车灯的光能或空调运行所需要的电能等。对于“汽油→发动机→车轮”这条能流而言,传递至车轮的能量越少,汽车获取一定动能所需要的汽油量就会越多,汽车的运行成本也将越高。如果成本过高,那么汽车可被选择作为交通工具的可能性将会降低。反之,传递至车轮的能量越多,汽车获取一定动能所需要的汽油量就会越少,汽车的运行成本也就越低,那么汽车可被选择作为交通工具的可能性就会增加。提高“汽油→发动机→车轮”这条能量传递单元能量传递效率的过程,实际上也是控制能量流动方向的过程。
伴随着以上这三个问题的解决,可以被我们选择作为交通工具的汽车也就应运而生,这个人也得以有机会选择驾驶汽车到达B地。虽然汽车的时速比自行车快很多,但仍然需要两天的时间才能到达B地。想到这,此人自言自语到:“如果我可以像孙悟空一样变出一个自己,然后让他替我去B地做事该有多好啊……”。
如果此人是第一位利用汽车作为交通工具去做事的人,那么当此人完成任务后,就会有越来越多的人乘坐汽车去做事,也就是说,汽车的应用得到推广。
透过这个事例,我们可以发现,对于一件不得不完成的事:
1)事情的完成过程往往需要消耗能量,如果此时存在其它有助于提高事情完成效率的能量,如上例中汽车的动能,那么,产生该能量的能流有可能会逐渐从独立于事情之外的事物变成该事情完成所需要的重要因素,甚至是不可或缺的因素;
2)当产生可借助能量的能流的稳定性得到保障时,该能流的应用价值才能得以体现;
3)产生可借助能量的能流的能量传递效率越高,该能流的应用价值越大,也就越容易得到推广;
4)如果多一个事情完成主体,也就是说,多一个做事的人,那么事情的完成效率也会得到进一步提升。
能流进化论认为,生物的出现是自然界中能量分布不均衡的结果,是自然界对能量传递单元选择的结果,是能量传递单元中能量传递介体功能不断升级的结果。正如汽车的诞生和演变是建立在“汽油→发动机→车轮”这一能流之上一样,非生命态的能流传递介体向生命体的演变过程也应是建立在一条相对稳定的能流之上的。那么,这条驱动生物出现的能流是什么?该能流又是如何驱动非生命态的能量传递介体一步步演变成生命体的呢?
(一) “能量可用系统”的出现
我们已经知道,任何生物都需要获取能量来维持其生命活动,说明一个可以最终演化为生命体的非生命体系,除了具有可介导能量传递的能力外,还应该具有完备的能量供给系统,用以维持其介导能量传递的能力。就像汽车的发动机,单纯的将其与汽油相连接并不能将汽油中的化学能转换为动能,还需要活塞的动能、点火系统的热能等其它能量的参与。那么,在非生命体系向生命体演变过程中,用于维持该体系能量传递能力的能量是什么?该能量又是如何被体系获取、储存和再分配的?
原子之间通过价键相互连接形成分子。价键(如氢键、酸酐键等)的断裂使得储存在物质分子中的化学能得以释放,而价键的形成也伴随着外界能量在物质分子中以化学能的形式存储(图1)。不同的化学物质含有不同类型或数量的价键,并且不同的价键储存不同的能量,因而,不同的化学物质含有不同的化学能。

图1 价键的合成与断裂伴随着能量的储存和释放
化学物质中,有的价键相对比较稳定,往往需要高温或高压才使其得以断裂或形成;有的价键则相对不稳定,容易受周边环境的影响。例如,磷酸化合物ATP是一个由一分子腺嘌呤、一分子核糖和三个相连的磷酸基团构成的核苷酸,分子简式A-P~P~P,式中的A表示腺苷,T表示三个,P代表高能磷酸基,“-”表示普通的化学键,“~”代表一种特殊的化学键,称为高能磷酸键。三个磷酸基团从腺苷开始被编为α、β和γ磷酸基,其中,α磷酸基团的酸酐键相对比较稳定,而β和γ磷酸基团的酸酐键则相对比较容易水解并释放出自由能。
价键断裂时,可释放出大量自由能的物质被称为高能化合物。这类物质在价键断裂过程中释放出的能量,有的足以驱动新的价键的形成或断裂,即驱动新的化学反应的发生。
研究发现,生物细胞内高能化合物的种类很多。根据它们键型的特点,可归纳为以下几种类型(注:此部分及下文中的化学反应式只是对所阐述内容的补充和解释,知道与否并不影响对所阐述内容的理解):1)磷氧键型,如乙酰磷酸、焦磷酸、磷酸烯醇式丙酮酸等;2)氮磷键型,如磷酸肌酸、磷酸精氨酸等;3)硫酯键型,如酰基辅酶A等;4)甲硫键型,如S-腺苷甲硫氨酸等。如果没有这些高能化合物,生物的生命活动将难以得到保障,生物也将不复存在。生物既然存在,表明这类物质在非生命体系向生命体的演变过程中,以及在维持体系能量传递能力方面起着重要作用。
这类含有相对比较容易断裂的化学键,同时,价键断裂时释放出来的能量,又能够驱动另外一种化学反应发生的高能化合物,被称为“能量可用系统” (图2)。

图2能量可用系统驱动化学反应的发生
我们知道,生命体是一个相对稳定的系统,说明其内化学反应的发生也是相对稳定的。这一特征间接表明,驱动化学反应发生的“能量可用系统”中的化学能是不会随意释放的,其可用性是一定条件下的可用性。如不然,储存在价键中的化学能就像会热能一样,只要存在能势差就可自发的释放和传递,那么,以其作为“能量可用系统”的生命体也将不会存在。一般情况下,“能量可用系统”中用于驱动化学反应发生的化学能需要在其它化学物质的催化作用下才能得以释放。例如,ATP可在ATP酶的催化作用下水解为ADP和磷酸根离子,同时释放出自由能。
化学反应的发生可能伴随着特定环境中新物质的出现。由于不同的化学物质储存着不同的化学能,因而化学反应具有两方面的环境意义:一是丰富环境中的物质种类多样性;二是驱动环境中的能量流动。第一点对于生命源起的意义在于,这些产物中,特别是新出现的物质,有些可能是构成生物结构的基本原料;第二点对于生命起源的意义在于,有些化学反应介导的能流可能是生命体构建的依托。
“能量可用系统”中的化学能在释放过程中,一部分转换为自由能,一部分转换为热能。热能用以维持体系的温度,进而使得体系中各种组分的活性及各种反应能够在一个相对稳定的温度范围内进行。自由能则主要用于驱动体系内各种化学反应的进行,为体系结构和功能的维持和延续提供保障。“能量可用系统”在释放能量的同时,自身也被转换成能量相对较低的物质。例如,ATP、GTP、NADH等“能量可用系统”释放的能量可驱动能量相对较低的丙酮酸转换为能量相对较高的葡萄糖,在这个过程中,这些“能量可用系统”也被转换成ADP、GDP、NAD+等能量相对较低的物质。这个过程可用下式表示(红色标记的化学物质属于“能量可用系统”):

无论是能量相对较高的物质被转换成能量相对较低物质,还是能量相对较低的物质被转化为能量相对较高物质,生命体内各种化学反应的发生都是有序进行的。这一特性(有序性)说明,生命体内各种化学反应的发生是外界环境对生命体做功的结果,这也说明了生物为什么会是自然界的产物。 基于“能量可用系统”能够驱动化学反应的发生可推知,“能量可用系统”具有两方面的自然价值:一是驱动环境中的物质转换;二是将相对难以在环境中流动的化学能转换成相对更容易流动的热能,更利于自然界实现其能量分布的相对均衡。由此可见,一个非生命体系所含“能量可用系统”的数量和种类越多,那么,该体系中发生的化学反应的种类就可能越多,体系的物质多样性就越丰富。已知构成一个功能细胞的化学物质种类繁多,表明拥有“能量可用系统”数量和种类越多的非生命体系,就越有可能演变成生命体。因此,拥有“能量可用系统”被认为是非生命体系可演变成生命体的第一要素,属于生命源起的第一阶段。
(二)“能量生产系统”的形成
“能量可用系统”可以提供驱动化学反应进行所需要的能量。然而,正如汽油中的化学能释放后,汽油将不复存在一样,随着能量的释放,“能量可用系统”也将不复存在,其功能也随之丧失。我们知道,生物均具有可维持其生命活动的属性,表明可最终演化为生命体的非生命体系,不但需要拥有“能量可用系统”,还应拥有使“能量可用系统”功能得到持续性保障的系统。那么,“能量可用系统”的保障系统是什么?这些保障系统又会具有什么样的特征?在回答这两个问题之前,我们先来看看下面两个众所周知的例子,看能否从中得到启发。
1)对于一些干旱地区,降雨量在年与年、季与季之间变化很大。然而,大多数用水,如农田灌溉、畜牧养殖、生活用水等都需要相对比较固定的用水量和用水时间,这就使得这些用水与天然降水在数量和时间上经常不能相适应。为了解决这个问题,往往会在这些干旱地区修建一些水库。在水库的选址上,往往还会优先选择有河流经过的地方,这样可以使水库里的水量容易得到保障。
2)对于汽车,要保障其汽油供应,不但需要在汽车中配备油箱,还需要有加油站、输油管道、汽油生产公司、石油开采公司等一系列的汽油保障系统。
透过这两个例子可以发现,要想保障“能量可用系统”功能的持续性,不但需要存在可以为其蓄能的,功能类似油箱或水库的“能量库”,还需要有保障“能量库”功能的能量供给系统。既然如此,那么可演化为生物的非生命体系中的“能量库”是什么呢?以及“能量库”中的能量又是如何得到保障的呢?
我们首先假设在一个特定的环境中,存在具有如下特点的两种化学反应:1)这两种化学反应中,一种属于合成反应,一种属于裂解反应;2)合成反应的产物恰好是裂解反应的反应物,同时,裂解反应的产物又是合成反应的反应物。此时,由这两种化学反应组成的体系,既是一种“物质循环系统”,又是一个开放系统(与周边环境存在物质和能量的交换)。在合成反应中,从周边环境输入该体系的能量被储存在新合成的化学物质中,而在裂解反应中,储存在反应物质中的能量,一部分会以化学能的形式储存在“能量可用系统”中,一部分会转换成热能、光能等相对更容易在环境中流动的能量,然后释放至环境中。
由合成反应和裂解反应组合而成的“物质循环系统”,在性质上类似可逆反应。二者主要差异在于,对于可逆反应而言,无论是正向反应还是逆向反应,均是由一步化学反应构成,而“物质循环系统”中的正向反应和逆向反应,均可由一步或多步化学反应构成。由此可见,可逆反应是“物质循环系统”中的一种特殊类型。
对于特定的可逆反应而言,反应物和产物在特定环境中的相对丰度可影响其化学反应的发生方向。也就是说,如果能量相对较高的化学物质在环境中的含量相对较高,那么该可逆反应的方向会倾向发生裂解反应的方向,反之,如果能量相对较低的化学物质在环境中的含量相对较高,那么可逆反应的方向会倾向发生合成反应的方向。同理,“物质循环系统”中各组分的相对丰度也决定了“物质循环系统”中的物质转化方向。前文说过,特定环境中的物质水平反映了其能量水平,物质分布反映了其能量分布。基于此可推知,“物质循环系统”也是一种能量传递介体,其化学反应的方向反映了环境中特定能流的流动方向,该方向是由该特定能流中的能量供体和能量受体在环境中的含量和分布情况决定的。换句话说,“物质循环系统”出现的根本原因是因为环境中的物质分布不均衡,或者说能量分布不均衡。
无论是生命体还是非生命体,在元素水平上都是一致的。例如,无论是人体中的氧元素,还是铁矿中的氧元素,它们的原子核都有8个带正电的质子,核外都有两个电子层,K层都有2个电子,L层都有6个电子等。然而,由这些元素组成的化学物质在生物体内却不断的发生变化,而且这些变化在生物体内所表现出来的多样性、普遍性和高转换频率在非生命体内是罕见的。由此推知,生命体与非生命物质统一于物理层面,分道于化学层面。这便意味着探寻分道的原因,也就是解析为什么有的体系中化学反应的发生是如此之多样、普遍和频繁,将有利于揭示生命的起源。自然界中,能量分布的不均衡性驱动了能流的产生,化学反应是能流的一种特殊呈现形式,“物质循环系统”既是一种特殊的化学反应类型,又是生物体的重要组成部分,这些再次说明,自然界中的能量分布不均衡性是生物出现根本原因。
对于一个拥有“物质循环系统”的体系而言,如果体系的能量水平高于周边环境,那么这个“物质循环系统”中的裂解反应将会被激活,在这个过程中:一部分化学能以热能的形式释放至周边环境中,一部分化学能储存至“能量可用系统”中。总之,裂解反应一方面降低了体系的总能量水平,保证了体系与周边环境之间的能量分布均衡性;另一方面使得体系自身的能量分布水平也相对更加的均衡(产物之间的能势差比反应前更低)。反之,如果体系的总能量水平低于周边环境的能量水平,那么这个“物质循环系统”中的合成反应部分将被激活,即利用从外界环境流入体系的能量以及体系中“能量可用系统”中的能量,将体系中能量相对较低的化学物质转化成能量相对较高的化学物质。在这个过程中,体系的总能量水平得以提升,同样保证了体系与周边环境之间的能量分布均衡性。由此可见,“物质循环系统”便是“能量可用系统”的“能量库”,其与所属体系之间的关系,就像油箱之于汽车,水库之于干旱地区。“物质循环系统”在体系中的价值在于维持体系与环境之间的能量均衡。为了便于与“能量可用系统”相对应,“物质循环系统”在后文中用“能量生产系统”表示(图3)。

图3能量生产系统是能量可用系统功能的保障
“能量生产系统”在所有细胞生物内普遍存在,可见其对生物的重要性。例如,由糖酵解途径和糖异生途径组成的“能量生产系统”(图4)。在糖酵解途径中,高能化合物葡萄糖经一系列的氧化分解过程,最终生成能量相对较低的丙酮酸,同时将价键断裂释放出的能量储存于ATP、NADH等“能量可用系统”中。在糖异生途径中,丙酮酸经一系列的酶促反应,利用储存在ATP、GTP等“能量可用系统”中的能量转变成葡萄糖。

图4 向下的箭头表示糖酵解途径中的化学反应,向上的箭头表示糖异生途径中的化学反应,绿色箭头表示只在糖酵解途径中的化学反应,棕黄色箭头表示只在糖异生途径中的化学反应,黑色箭头表示两个途径都有的化学反应,属于可逆反应。
当细胞所在环境中的葡萄糖含量充足时,即环境中的能量高于体系(细胞)中的能量,细胞中的糖酵解途径被激活,整个过程可用下式表示(红色标记的化学物质属于“能量可用系统”):

如果细胞所在环境中的葡萄糖含量匮乏,即环境中的能量低于体系(细胞)中的能量,丙酮酸会经糖异生途径重新合成葡萄糖。这个过程可用下式表示(红色标记的化学物质属于“能量可用系统”):

同理,脂肪酸的合成代谢和分解代谢、氨基酸的合成代谢和分解代谢、核苷酸的合成代谢和分解代谢等组合而成的系统,均属于“能量生产系统”。这类“能量生产系统”的合成反应部分可以仅靠体系自身的“能量可用系统”得以实现。
除此之外,还有一类“能量生产系统”,它们需要外界能量的参与才可以正常运行。例如,在光合自养型生物体中普遍存在的,由光合作用和呼吸作用组成的“能量生产系统”。光合作用是指绿色植物利用叶绿素等光合色素或者某些细菌(如带紫膜的嗜盐古菌)利用其细胞本身,在可见光的照射下,将二氧化碳和水(细菌为硫化氢和水)转化为葡萄糖等有机物,并释放出氧气(细菌释放氢气)的生化过程。在这个过程中,光能被转换成化学能储存在葡萄糖等有机物中。呼吸作用则是指细胞中的葡萄糖等有机物,经过一系列的氧化分解反应,变成二氧化碳和水,同时释放出能量,以维持生物生命活动需要的生化反应过程。在这个过程中,葡萄糖等有机物裂解释放出的能量,一部分以热能的形式释放出来,一部分被储存在ATP、NADH等“能量可用系统”中。
综上所述,“能量生产系统”的出现,既驱动了体系自身能量的流动,又驱动了体系与周边环境之间的能量流动;既维持了体系自身的能量水平,又实现了体系与环境之间能量分布的相对均衡;既是“能量可用系统”功能稳定性的保障,又是非生命体系演变成生命体的必要因素。已知发生在一个功能细胞内部的化学反应种类繁多,因而推知,拥有“能量生产系统”数量和种类越多的非生命体系,其物质水平和能量水平就越稳定,功能也就越容易得到保障,也就越有可能演变成生物体。因此,拥有“能量生产系统”并实现其与“能量可用系统”的统一,被认为是非生命体系向生物演变进程中的第二个阶段。
(三)“竞争”和“合作”
(1)“能量可用系统”功能的限定
我们知道,细胞内同一种“能量可用系统”释放出的能量,可以驱动多种化学反应的发生。例如,储存在“能量可用系统”ATP中的自由能,可以催化以下几种化学反应的发生:

……
对于非生命体系中的“能量可用系统”,其驱动的化学反应中,有的可能会促进非生命体系向生命体的演变,有的可能会抑制这一过程的演变,也有的可能与这一演变过程毫无关系。这就意味着,如果“能量可用系统”中的能量随机释放、随机应用,那么这些能量驱动非生命体系最终演变成生命体的概率,就好比给猴子一台打字机,让它通过随机敲打并最终写出一部《红楼梦》一样。既然生物已经在自然界中出现,说明可演变为生物的非生命体系中的“能量可用系统”,其能量的释放和应用过程不可能是随机的。那么,究竟是什么力量限定了“能量可用系统”中能量的应用范围,以至于这部分能量主要用于驱动生命源起的呢?
在我们身边,拥有“能量可用系统”和“能量生产系统”的非生命体系的种类非常多,而且现在也可以很容易人为制造,如智能手机、智能机器人、电子手表、汽车等。然而,我们并没有从这些系统中看到它们有演变成生物体的迹象。那么,最终可演变成生物的非生命体系究竟还需要具有什么样的特性呢?
(2)能量传递单元中的“共享物质”
生物细胞内含有多种多样的能量传递单元,例如,

……
这种现象表明,可最终演化为生物的非生命体系中,必然存在种类繁多的能量传递单元。在这些能量传递单元中,不同的能量传递单元之间难免会出现交集。也就是说,几种能量传递单元共享同一种能量供体、能量受体或者能量传递介体。这种被几种能量传递单元共享的物质,称之为“共享物质”。依据在能流中的角色,“共享物质”可分为以下三种:
1)共享能量供体。当“共享物质”是能量供体时,称之为“共享能量供体”,这类物质存在多种不同的能量传递途径。同一种“共享能量供体”在一个体系内可能存在多个拷贝,不同的拷贝可能参与不同的能量传递途径。然而,对于一个特定的“共享能量供体”而言,无论它有多少种能量传递途径可以选择,它只能选择其中一种。这是因为,对于某个特定的“共享能量供体”,当其能量释放时,价键就会发生断裂,此时,产物就已经不再是原来的物质分子。
2)共享能量受体。当“共享物质”是能量受体时,称之为“共享能量受体”,这类物质可接受来自多条能量传递途径的能量。同一种“共享能量受体”在一个体系内可能存在多个拷贝,且不同的拷贝参与不同的能量传递单元。然而,对于一个特定的“共享能量受体”而言,无论它有多少种能量来源可供选择,它只能选择其中一种。这是因为,对于某个特定的“共享能量受体”而言,当它接受能量并转变成能量相对较高的物质分子时,新生成的物质分子同样不再是原来的物质分子。
3)共享能量传递介体。当“共享物质”是能量传递介体时,称之为“共享能量传递介体”,这类物质可以介导不同种类的能量供体和能量受体之间的能量传递。对于“共享能量传递介体”而言,无论它可以与多少种能量供体和能量受体组成能量传递单元,对于一个特定的能量传递介体,它所介导的能量流通量是有限的。就像任意一条河流,其河道的宽度和深度决定了该河流传递水的能力,当水流量超过了河道的传递能力时,多余的水只能经其它河道传递。
“共享物质”在能量传递单元中所表现出来的特征在生物介导的能量传递单元中也同样普遍存在。
1)共享能量供体:例如,对于羚羊而言,狼和狮子都以其作为能量供体。也就是说,羚羊是狼和狮子的“共享能量供体”。然而,对于一只羚羊而言,它要么被狼吃掉,要么被狮子吃掉,而不可能同时被狼和狮子吃掉。即使是几头狮子共享一只羚羊,被其中一头狮子吃掉的部分也不可能同时被另一头狮子吃掉。
2)共享能量受体:例如,氧气既是狼的能量受体,又是狮子的能量受体,也就是说,氧气既可以在狼的细胞内接受代谢产生的电子变成水,又可以在狮子的细胞内接受代谢产生的电子变成水。然而,对于一个氧气分子而言,当它在狮子的细胞内变成水后,就不可能再在狼的细胞内行使同样的功能。
3)共享能量传递介体:对于生活在同一片草原上的兔子、羚羊和狮子而言,兔子和羚羊都可作为狮子的能量供体,因而狮子是兔子和羚羊的共享能量传递介体。一般情况下,当狮子在吃羚羊的时候,不会再同时去吃兔子。
在生物界存在一个有趣的现象,大多数的生物都可以利用多种能量供体,即可以以多种物质作为食物。然而,每一种生物都有其相对更喜好的食物。当生物作为能量传递介体时,该生物最喜好的食物越充足,它所变现出来的食物专一性便越显著。例如,大肠杆菌既可以利用葡萄糖作为能量供体,也可以利用乳糖作为能量供体。当培养环境中同时存在葡萄糖和乳糖时,大肠杆菌会优先利用葡萄糖,只有葡萄糖消耗殆尽后,才会以乳糖作为能量供体。当培养板中的葡萄糖含量充足时,对于观察者而言,大肠杆菌所表现出来的现象便是葡萄糖是其专一的能量供体。这说明,共享能量传递介体的共享性的呈现与否与环境中能量供体或能量受体的种类和丰度相关。
(3)能量传递单元的专一性和高效性
在一个特定的体系中,物质分子的布朗运动,以及与环境中其它物种分子之间的相互作用,使得含有“共享能量传递介体”的不同能量传递单元之间的比例时刻发生着变化,而且这种变化具有随机性,这便使得特定环境中含有“共享能量传递介体”的能量传递单元具有不稳定性。这一不稳定性进而导致所属体系结构和功能的不稳定。细胞生物学的发展已经表明,生物细胞的结构和功能是相对稳定的,这就说明,一个结构和功能不稳定的非生命体系是不可能演化成生物体的。反过来讲,生物的出现说明,“共享能量传递介体”的“共享”特性在生命源起的过程中被逐渐弱化,甚至被功能专一的能量传递介体所取代,结果便是,共享能量传递介体逐渐演变为只对应特定能量供体和能量受体的能量传递介体。
尽管功能专一的能量传递介体使得其所属能量传递单元的结构组成稳定性得到保障,然而,其与能量供体和能量受体组成完整能量传递单元的几率却降低。例如,狼既可以吃羚羊,又可以吃兔子。假设现在在狼群中出现了一匹变异了的狼,该狼只吃兔子且捕获兔子的能力与正常的狼相同。可想而知,变异狼的功能专一性更强,但遇到食物的几率要比正常的狼更低,被饿死的几率也就随之增加,进而表现为能量传递能力降低。该变异狼要想与正常狼具有相似的能量传递能力,唯一的途径便是提升其捕获兔子的能力。当捕获兔子的能力得到提升后,该变异狼被饿死的几率便会降低,进而表现为能量传递能力增强。然而,捕获兔子能力的提升,意味着兔子可能会被更快的吃光,进而以兔子作为唯一食物的变异狼被饿死的几率随之增加。也就是说,随着时间的推移,变异狼有可能又会表现为能量传递能力降低。为与“专一性”相对应,我们用能量传递效率来表征能量传递能力。那么,当能量传递能提升后,能量传递介体的能量传递效率增加,即表现为“高效性”。能量传递介体能量传递能力的高效性,增强了所属能量传递单元的能量平衡能力,但随着时间的推移,能量传递单元的结构组成不稳定性也有可能随之增加。
综上所述,能量传递介体的专一性使得其所属能量传递单元的结构组成相对更加稳定,但能量平衡能力减弱;能量传递介体的高效性使得其所属能量传递单元的能量平衡能力增强,但结构组成更加脆弱。能量传递介体的高效性和专一性相互影响、相互制约,二者的统一使得该能量传递介体所属能量传递单元的能量平衡能力得以更好的适应环境的需求,也就有存在下去的可能。这也解释了为什么细胞中的各种催化酶类能量传递介体具有高效、专一的特性。总之,最终在生命的源起过程中保留下来的能量传递介体是高效性和专一性实现统一的、相对更加适合环境要求的能量传递介体。
(4)“共享物质”对能量传递单元的选择性
假设能量传递单元“S→I→C”和“S→II→D”共存于一个体系中(图5),“S”为这两种能量传递单元的“共享能量供体”。从“S”的角度讲,它所储存的能量有两种能量传递单元可供选择。反过来,从能量传递单元的角度看,这两种能量传递单元对“S”中的能量表现为“竞争”关系。“S”中的能量最终流经的能量传递单元被认为是这场“竞争”中的胜者,而没有流经的能量传递单元则被认为是这场“竞争”中的败者。
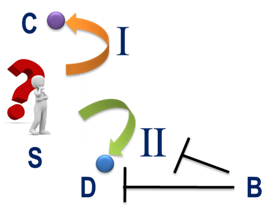
图5能量的传递途径具有选择性。“S”为能量传递单元“S→I→C”和“S→II→D”的“共享能量供体”,“I”和“II”为两种能量传递介体,“B”为“D”或“II”的抑制因子。
假设能量传递介体“I”和能量传递介体“II”的能量传递能力相同。如果“S”与能量受体“C”之间的能势差比“S”与能量受体“D”之间的能势差更利于“S”中的能量的传递,那么“S→I→C”所表现出来的能量平衡能力要比“S→II→D”更强。在恒温、恒压条件下,存在能势差的两种化学物质之间不一定就会发生能量传递,只有存在合适的能势差才有发生能量传递可能。例如,葡萄糖和1,3-二磷酸甘油酸两种物质分子均与ADP均存在能势差,然而,一般情况下,葡萄糖不会直接将能量传递给ADP,而是先经过一系列的转换变成1,3-二磷酸甘油酸后,才将释放出的能量储存入ADP,使其转变成“能量可用系统”ATP。此时,可以认为,1,3-二磷酸甘油酸与ADP之间的能势差比葡萄糖与ADP之间的能势差更适合能量传递。生物细胞内不同物质分子之间均存在能势差,但各种生化反应的发生井然有序,能量在其中的传递有条不紊,说明合适的能势差是化学物质之间能量传递的必要条件。一般情况下,两种物质之间的能势差越合适,能量传递越容易发生,传递效率也会越高。
假设“S”与能量受体“C”之间的能势差和“S”与能量受体“D”之间的能势差相同,如果能量传递介体“I”比能量传递介体“II”的能量传递能力更强,根据前文所述,“S”中储存的能量同样更倾向于沿能量传递单元“S→I→C”进行传递。 如果存在物质分子“B”,其可通过与能量传递单元“S→II→D”中的分子“D”或“II”相互作用,抑制“S”中的能量向“D”传递,那么“S”中的能量同样倾向于沿能量传递单元“S→I→C”传递。此时,物质分子“B”与能量传递单元“S→I→C”之间可以看作是一种“合作”关系,“B”作为“S→I→C”的环境因素,参与了能量传递单元“S→I→C”与能量传递单元“S→II→D”之间对“S”中能量的争夺。
综上所述,对于“共享能量供体”而言,其能量在传递时有两种因素对传递途径的选择起重要调控作用:一种是可选择的能量传递单元的能量传递效率,主要与能量传递介体和能量受体的性质有关;另一种是环境因素,也就是能量传递单元以外的物质分子对能量传递单元能量传递效率的影响。
(5)“竞争”和“合作”
“竞争”与“合作”是可以介导同种能量流动的物质分子之间所表现出来的关系,即理论上可介导能量流动的物质分子与实际介导能量流动的物质分子之间的关系。实际介导能量流动的物质分子之间表现为“合作”关系,而理论上可以介导能量传递但实际中并没有参与能量流动的物质分子,与实际介导能量流动的物质分子之间表现为“竞争”关系。能量传递单元中“共享物质”的存在是“竞争”和“合作”出现的根本原因。小至不同的物质分子,大至不同的生物或由生物组成的群体,都可以作为“竞争”或“合作”的对象。
1)物质分子作为“竞争”和“合作”对象。
假设存在两株细菌,它们均以葡萄糖作为唯一能量供体。现在,我们在添加葡萄糖的培养板中培养这两株细菌。在这个体系中,葡萄糖是这两株细菌的“共享能量供体”,而这两株细菌则是葡萄糖可选择的能量传递介体。此时,这两株细菌之间就存在对葡萄糖的“竞争”关系。葡萄糖的含量越少,二者所表现出来的“竞争”就越显著。
对于这两株细菌,假设其中一株具有青霉素抗性,即可以在添加青霉素的培养板中生长,而另一株则是青霉素敏感型,即青霉素可以抑制其生长。如果在添加葡萄糖的培养板中添加青霉素,然后将这两株细菌接种至该培养板中,就会发现,青霉素抗性菌株可以正常生长,而青霉素敏感性菌株不能正常生长。在这个例子中,“青霉素”与“青霉素抗性菌株”之间便是一种“合作”关系。
2)“能量生产系统”作为“竞争”和“合作”对象。
“能量生产系统”一般由多种物质分子组成,不同的“能量生产系统”含有不同的物质组成,行使的功能也有所差异,甚至供应不同种类的“能量可用系统”。前文讲过,“能量可用系统”是体系结构和功能得以发挥的保障,而“能量生产系统”是“能量可用系统”功能持续性的保障。为了保障“能量可用系统”功能的稳定,“能量生产系统”需要维持其能量水平。当流入一个体系的能量有限时,体系中的不同“能量生产系统”就可能会对这些能量进行“竞争”。“能量生产系统”为了增强自身的竞争能力,就需要自身不同组分之间,或者与该“能量生产系统”存在“利益”关系的组分之间加强“合作”。例如,现在有两个“能量生产系统”,一个是由“脂肪酸合成代谢途径”和“脂肪酸降解代谢途径”组成的“能量生产系统”;一个是由“糖酵解途径”和“糖异生途径”组成的“能量生产系统”。当细胞所在环境中的葡萄糖含量比较低时,这两种“能量生产系统”之间便存在着“合作”关系,即脂肪酸经降解代谢转变成葡萄糖,葡萄糖在糖酵解途径被进一步裂解,最终转变成丙酮酸。这个过程释放出的自由能储存至ATP等“能量可用系统”中,用以维持细胞结构和功能的稳定。对于这个过程,我们可以认为,由于这两种“能量生产系统”之间的“合作”,细胞的结构和功能才得以在葡萄糖含量有限的环境中得到保障。
当环境中的葡萄糖含量相对比较高时,细胞内的糖酵解途径和脂肪酸合成代谢过程均需要葡萄糖的参与。此时,可以认为这两种“能量生产系统”之间存在对葡萄糖的“竞争”,也就是说,此时,这两种“能量生产系统”之间是“竞争”关系。
3)生物作为“竞争”和“合作”对象。
对能量的“竞争”和“合作”在同一个生物的不同组织、器官之间,不同生物之间,甚至不同物种之间普遍存在。这是因为,生物结构和功能的维持都需要消耗能量。例如,已知一群狼与几头狮子均盯上了它们的“共享能量供体”羚羊。对于正在捕食的狮子而言,它的爪子和牙齿之间是一种“合作”关系;若这几头狮子都参与了捕食过程,那么这几头狮子之间也是一种“合作”关系;若狼吃了狮子没有吃完的羚羊肉,对于储存在羚羊中的能量而言,狼与狮子也是一种“合作”关系;为了促进吞入肚子中的储存在羚羊肉中的能量释放,狮子自身的组织细胞和其肠道中的肠道微生物之间也是一种“合作”关系……
如果羚羊的数量有限,狼和狮子之间为了自身的生存就需要对羚羊展开“竞争”;如果羚羊的数量仅够维持一头狮子的存活,那么这几头狮子之间也会存在对羚羊的“竞争”;如果羚羊的数量难以维持一头狮子的存活,那么狮子与自身肠道里的微生物之间也会存在能量的“竞争”;肠道里的不同微生物之间同样也会存在对进入肠道中的能量的“竞争”……
(6)“竞争”和“合作”驱动的分工
对于处于同一个体系中的物质而言,按其在所属体系与其它体系进行能量“竞争”过程中的分工,可分为两类:一类是中央能量控制系统;一类是辅助系统。按照物质分子在所属体系与其它体系进行能量“竞争”中所起的作用,辅助系统又被分为攻击系统和防御系统两类(图6)。
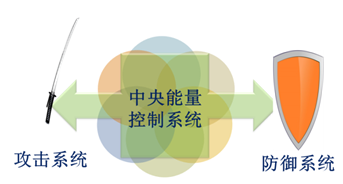
图6一个体系可分为中央能量控制系统和辅助系统两类,辅助系统又可进一步分为攻击系统和防御系统两类
1)中央能量控制系统。主要负责能量的获取、传递、储存、分配等。该系统可有效提高一个体系的能量传递及利用效率,在介导能量供体中的能量进行释放和传递的同时,也维持着自身结构和功能的稳定。在细胞内,参与糖酵解途径、三羧酸循环、磷酸戊糖途径、电子传递链、脂肪酸代谢等过程的物质分子都属于这个系统,它们在能量的释放和传递过程中发挥着重要作用。此外,ADP、GDP、NAD+、FAD等能量受体和ATP、GTP、NADH、FADH2等“能量可用系统”在能量储存和分配过程发挥着重要作用。此外,该系统中的能流也是非生命体系向生命体演变过程中所依附的“地基”。
2)辅助系统。辅助系统中的攻击系统主要用于抑制或削弱其竞争者的竞争能力,而防御系统则主要用于维持自身系统的稳定性以及抵抗来自竞争者的攻击。例如,肺炎球菌、炭疽杆菌等在细胞外层具有黏性或胶状的荚膜,有了荚膜的保护,这类细菌就好像穿上了防弹衣,可以抵抗人体吞噬细胞的吞噬作用,还可以有效保护细菌免受或少受多种杀菌或抑菌物质(如溶菌酶等)对细菌的损伤。由此可见,荚膜是细菌的一种防御系统。
破伤风梭菌是一种来自土壤的厌氧菌,它分泌的破伤风毒素对人的中枢神经系统有高度的亲和力。此毒素一旦进入中枢神经系统,便会固定在神经突触上影响甘氨酸的释放,进而引发肌肉僵直痉挛、抽搐和瘫痪。此例中,破伤风毒素便是破伤风梭菌产生的攻击武器,属于攻击系统。
总之,辅助系统的出现,使得所属体系的能量传递能力的稳定性和持续性得到保障,进一步驱动了非生命体系向生物体的转变。
一个“共享物质”一次只能选择一种能量传递单元。能量传递的这种唯一性加剧了竞争者之间的竞争,也推动了竞争者之间的“军备竞赛”。此外,竞争者的组成越复杂,各组分之间所表现出来的“合作”就越显著,竞争力也就越强。
既然生物的价值在于驱动自然界中的能量分布实现相对均衡,那么,为了更有效的实现自身的价值,可演变成生物的非生命体系中所含有的“共享物质”所选择的能量传递途径,在很大程度上,也应该是最有利于所属非生命体系发挥功能的那条途径。根据前文所述,自然界中的能量均衡程度决定了自然界所需要的能量平衡能力,进而决定了能量传递介体的能量传递能力。由此可推知,自然界对体系能量传递能力的要求,在大方向上制约了体系中的“共享物质”对体系中能量传递单元的选择。“竞争”和“合作”是自然界在选择其最适能量传递单元过程中所呈现出来的一种表现形式,是自然界选择的表现,二者既限定了体系的演变方向,又驱动了体系的演变。因此,“竞争”和“合作”的出现以及其与非生命体系演变方向的统一,被认为是非生命体系向生物体演变进程中的第三个阶段。
(四) 遗传系统的出现及生物的诞生
一般情况下,一个体系的能量竞争能力越强,其能量传递能力也就越强,从环境中获取的能量也就越多。由于一个体系结构和功能的维持需要消耗能量,对于从环境中获取能量越多的体系,其结构和功能得到升级的几率就越高,也就是说,越利于自己的演变。从环境对能量平衡能力需求的角度而言,竞争能力越强的体系,也就越适合环境的需求。对于竞争过程中的失败者而言,由于难以通过获取足够的能量来提升自身的能量传递能力,致使其所属能量传递单元的能量平衡能力越来越弱,也就越来越难以适应环境的需要。随着时间的推移,优胜者与失败者之间的差距越来越大。由此可见,生物在进化过程中所呈现出来的“物竞天择、适者生存”这一现象,在生命源起过程中就已经开始呈现。
在开头所述的故事中,一个要去B地出差的人可以通过改进出行交通工具来缩短出行时间。当交通工具难以得到进一步提升时,最好的可能便是在B地有一个与此人一样的人来替他完成需要在B地做的工作。对于一个能量分布不均衡的环境而言也是如此。当优胜者的个体能力在一定时期内难以得到进一步提升时,若要环境中的能量分布得以更有效的实现均衡,最有效的措施便是在环境中再产生一个与当前竞争优胜者一样的体系。事实表明,所有的生物物种都具有繁殖的能力,那么这个能力是如何在自然界的能量流动过程中产生的呢?
在外界环境因素(如水流、风等)的影响下,对于特定环境中的优胜者而言,不仅自身的结构组成是动态变化的,整个体系也有可能会被转移至另外一种环境。在这种情况下,昨日的优胜者不一定是今日的优胜者,此处的优胜者也不一定是彼处的优胜者。对于不同环境中的优胜者,有些拥有相对稳定的组分,这些组分不会因优胜者在环境中的转移或环境的变化而发生显著的改变,它们通过相互协作,能够利用新的环境中的物质分子,重新组合成与原来一样的优胜者。也就是说,这些稳定的组分可以使优胜者在新的环境中得以复制。
此处可能有人会问,这些稳定的组分具体是什么?可以明确的说:“无法回答”。这是因为,如果谈具体的组分,便涉及到具体的物质分子,对生物学的探索已经证明:1)与生物遗传有关的物质分子种类繁多,如各种DNA、RNA和调控因子等;2)虽然有些遗传元件,如DNA聚合酶、RNA聚合酶等在不同的物种内有一定的序列同源性,但还是有很多的物质分子存在差异,如真核生物的基因中含有内含子,而原核生物基因中不存在内含子等;3)同一种遗传元件可能有多种生物学功能,而且在不同的物种中行使的功能也会有差异,如DNA聚合酶不止有合成DNA的功能,还有修复DNA的功能等;4)各种遗传元件之间的联系复杂多变……这些现象表明,从物质分子的角度去探索生命的源起几乎是难以完成的工作,这部分在前文也已有阐述。这也是为什么《隐藏的动力:生物在自然界中的价值》一书要从所有生物都呈现出来的,在驱动自然界中能量流动方面的价值的角度来解析生命源起的原因。因此,该书不关心这些遗传组分是什么,只是根据已知生物的功能,去推断所拥有的组分应该具有什么样的功能,尤其在能量流动中的功能,或着说在生物价值实现中的功能。
既能在优胜者中稳定存在,又能使优胜者的功能延续性得到保障的系统,即利于优胜者在环境中再产生一个与优胜者功能相同或相似的系统,称之为“遗传系统”(图7)。
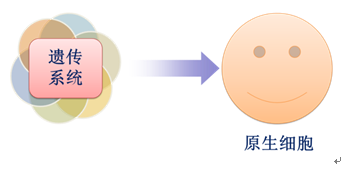
图7 遗传系统的出现
遗传系统的出现,不但使体系在环境中的能量传递能力得到进一步提升,还使得这种能量传递能力得以延续。因此,遗传系统的出现,被认为是非生命体系演变成生物进程中的第四个阶段,而拥有遗传系统的体系,便是现在生物结构和功能的基本单元(细胞)的雏形,即原生细胞。
(五) 小结
综上所述,能量守恒定律和热力学第二定律驱动的自然界中的能量流动是非生命体系向生物演变的驱动力,自然界中的能量分布不均衡性是非生命体系向生物演变的根本原因。作者认为,能量传递介体从非生命态体系向生物演变的过程主要依次经历以下四个阶段:1)拥有“能量可用系统”,用以提供非生命体系向生物演化过程中所需要的能量;2)拥有“能量生产系统”,用以保障“能量可用系统”功能的持续性;3)能量会以当前最高效的能量传递方式传递能量这一特征驱动“竞争”与“合作”这一现象的出现,并因此限定了非生命体系的演变方向;4)遗传系统的形成,使得特定体系的能量传递能力得到延续。
https://blog.sciencenet.cn/blog-3479210-1309538.html
上一篇:能流进化论——自然选择下的生命演变三部曲
下一篇:能流进化论——生物进化
全部作者的精选博文
- • 再谈科研思路
- • 科研的思路应该是什么?
- • 面点统一的高等教育培养模式
- • 我眼中的王老师
全部作者的其他最新博文
- • 冲破枷锁不容易
- • 再谈科研思路
- • 科研的思路应该是什么?
- • 再读吉布森的生态光学
- • 赋予冰冷的数字以生物学意义
- • 大道至简