博文
【重磅】基因编辑“女神”Jennifer等发现新CRISPR-Cas系统!
 精选
精选
||
本文首发于微信公众平台“科研小助手”,ID:SciRes。微信扫描文末二维码添加关注!

今日凌晨,Nature杂志Online了一篇题为“New CRISPR–Cas systems from uncultivated microbes(来自不可培养微生物的新CRISPR-Cas系统)”的文章,该文主要报道了两种新的CRISPR-Cas系统:CRISPR–CasX 和CRISPR–CasY。该文章的通讯作者是加州大学伯克利分校Jennifer A. Doudna和Jillian F. Banfield。有意思的是,在文章中特别注明了该项发现已递交临时专利申请(provisional patent application),其中三位共同一作以及两位通讯作者均名列发明者一栏,在这件事情上是由于prior-art原因还是吸取了与张峰专利之争的教训,耐人寻味。
而Nature更是火速帮其Online了这篇文章(Accelerated Article Preview),更有趣的是几乎在该文Online的同时,包括澎湃新闻等国内外媒体就放出了该文的相关报道!

文章详解
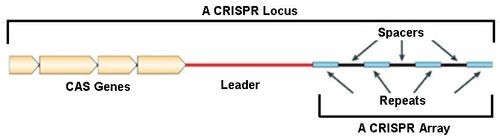
CRISPR-Cas系统为微生物提供适应性免疫。CRISPR是位于细菌和古生菌基因组的特殊的DNA重复序列,通常由一个前导区(Leader),多个短而高度保守的重复序列区(Repeat)和多个间隔区(Spacer)组成。前导区富含AT长度为300~500bp的区域,是CRISPR的启动子序列。重复序列区长度为21~48bp,具有回文序列,可形成发卡状二级结构。重复序列之间被长度为26~72bp的间隔区隔开。间隔区由细菌或古生菌俘获外源DNA组成,类似免疫记忆,当含有同样序列的外源DNA入侵时,可被细菌机体识别,并进行剪切使之表达沉默,达到保护自身安全的目的。
对CRISPR位点侧翼序列分析发现,在其附近存在一个多态性家族基因。该家族编码的蛋白质均含有可与核酸发生作用的功能域(具有核酸酶、解旋酶、整合酶和聚合酶等活性),并且与CRISPR区域共同发挥作用,被命名为CRISPR关联基因(CRISPR associated),缩写为Cas。目前发现的Cas包括Cas1~Cas10等多种类型。Cas基因与CRISPR共同进化,共同构成一个高度保守的系统。(如下图)
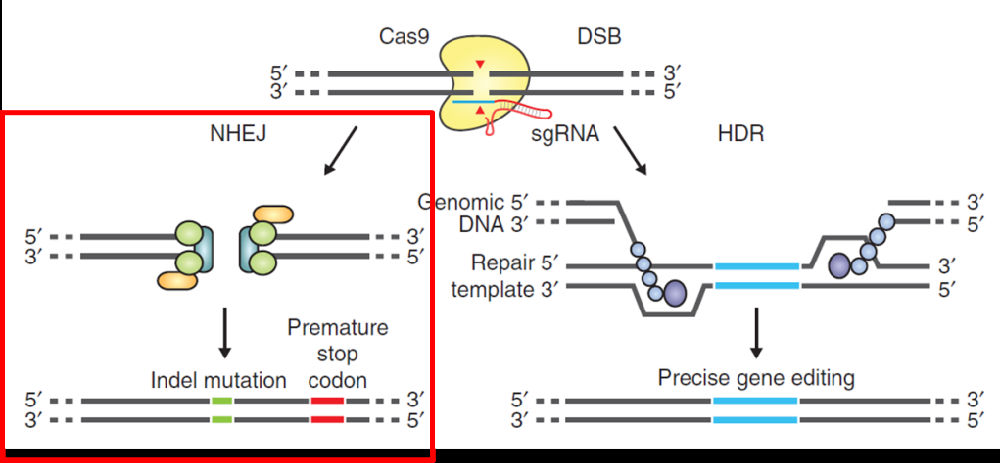
目前发现的 CRISPR-Cas系统主要分为三种类型,研究得最为详细的是第二种类型。II型CRISPR-Cas系统是由一个Cas蛋白结合向导性RNA(Guide RNA)到特异的位点,然后切割序列,造成DNA损伤。通常两种方式修复:一种是易错的修复机制(error prone repair),往往会造成碱基的插入和缺失(indel)另外一种是模板介导同源重组修复(HDR)。(如下图)
CRISPR-Cas系统的特点也为物种改良和很多遗传性和肿瘤等疾病带来革命性的改变。然而现有的这些CRISPR-Cas系统是根据分离培养的细菌体内发现的。众多的环境微生物中CRISPR-Cas系统还是没有被研究过。宏基因组学是研究环境微生物基因组的分支学科,这样为诠释环境微生物的基因功能提供可能。加州大学伯克利分校的Jillian F. Banfield教授领导的研究小组利用宏基因组研究技术从环境微生物发现了更多的新的CRISPR-Cas系统。包括从纳古生菌中发现了Cas9蛋白,这也是第一次在古生菌种找到Cas9蛋白,跟以前发现的Cas9并不相同。在细菌中,发现了两种新的CRISPR-Cas系统:CRISPR-CasX和CRISPR-CasY系统。这些新的CRISPR-Cas系统拓宽了识别DNA序列的范围,为发展CRISPR-Cas系统的应用于生物技术领域和疾病治疗等提供更多的可能和更多的选择。
Jillian F. Banfield教授领导的研究小组从地下水,沉积物,矿山酸性废水,土壤,婴儿肠道等已经测序入库的T碱基的数据库中寻找II型CRISPR-Cas系统。
通过生物信息学分析,寻找跟CRIPSP簇和Cas1序列类似的基因位点。分析了数据库中大约1.5亿个蛋白编码基因。他们第一个发现了古生菌中的Cas9蛋白,又从细菌中发现了CRISPR-CasX和CRISPR-CasY。他们结构特点如下图。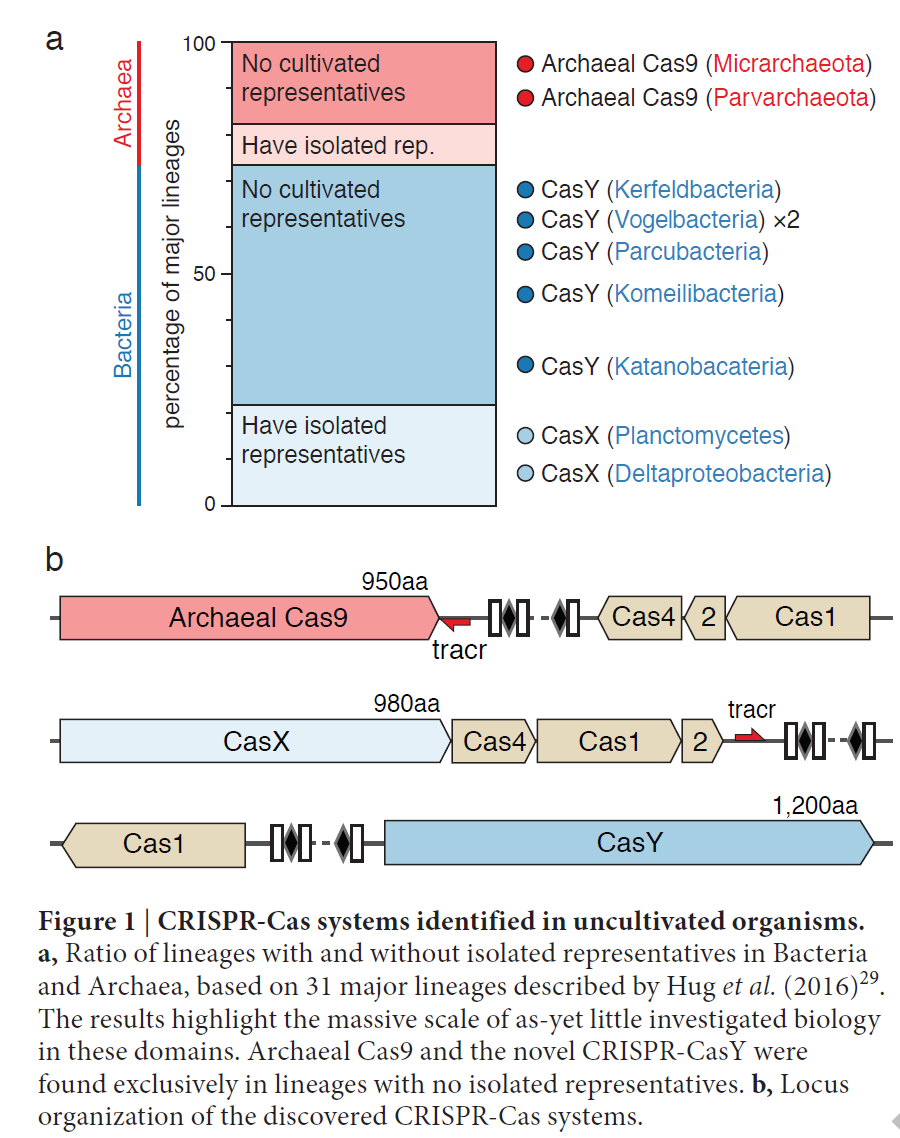
古生菌ARMAN-1的CRISPR-Cas 位点包括很大的 CRISPR基因簇和相连的Cas1, Cas2, Cas4和Cas9。作者分析了16份古生菌样本,CRISPR-Cas位点的序列同源性高达99.7%。对古生菌ARMAN-1其它的CRISPR-Cas序列分析后发现它们具有保守的末端和中间的可变的区域,表明插入了不同的序列(图4a)。作者推测位于Cas9基因和可变区域中间的200bp, 是启动子和转录起始点。间隔区的高可变性表明古生菌ARMAN-1中的CRISPR-Cas9是有活性的。系统进化分析Cas1序列表明古生菌ARMAN-1的CRISPR-Cas系统不同于已经发现的II型CRISPR-Cas系统其它成员。Cas4相关的CRISPR-Cas系统属于II型B,而Cas9序列跟II型C系统更类似。他们发现古生菌的II型系统融合了B和C的特点。
56个古生菌ARMAN-1间隔序列靶向10Kb的环形序列,主要是编码很多短的预测的蛋白序列,这个10Kb的环形序列很多能就是古生菌的病毒序列。有研究用冷冻电镜技术观察到附着在古生菌上病毒颗粒。ARMAN-1的间隔序列也来源于ARMAN-2和Thermoplasmatales古生菌转座子序列。有研究发现了ARMAN 和Thermoplasmatales细菌接合的桥联结构,说明这两种古生菌之间存在某种联系。
因此说ARMAN-1的CRISPR-Cas9系统是防止外来转座子,维护基因组的稳定性,类似于真核生物生殖细胞中的piRNA防止生殖细胞转座的作用。
ARMAN-4的Cas9基因只有一个CRISPR-间隔序列基元。也不是ARMAN-1那样还存在其它 Cas蛋白。也没有发现它们的间隔区域靶向的位点。但是在收集的样品中这个位点还是很保守的,它是一个单靶位点的CRISPR-Cas系统。它们有不一样的生理作用,如基因调控和细胞间的相互作用。
CRISPR-Cas系统在PAM序列旁边的2-4个和苷酸序列区分自我和非我。图2c 所示,ARMAN-1的基因组上PAM序列旁边更偏好为NGG,Cas9和crRNA和tracrRNA一起靶向到特异位点切割。前面认为古生菌没有II型CRISPR系统是由于它没有RNA聚合酶III,这个是负责crRNA/tracrRNA复合体的成熟。在ARMAN-1的基因组中没有发现RNA聚合酶III,在 CRISPR位点也没有发现启动子结构,说明ARMAN-1通过Guide RNA成熟有其它机制,有待阐明。大肠杆菌和酵母中表达纯化的古生菌ARMAN-1和ARMAN-4的Cas9蛋白的体外实验,未见它们有切割能力,说明了古生菌独特的系统,很可能它们在蛋白翻译后修饰很不一样,造成外源表达的Cas9不具有活性。
除了Cas9以外,II型系统还有Cpf1,C2c1和C2c2。另外一个基因C2c3存在并不广泛。作者假设在环境微生物中可能还存在着其它的Cas蛋白酶。通过宏基因组数据库比对,作者在地下水和沉积物样本中找到了一个新的 Cas蛋白酶,大约为980个氨基酸序列长,命名为 CasX。Phyla,Deltaproteobacteria和Planctomycetes种属的细菌中CasX同源性达到68%。序列比对发现CasX蛋白酶跟转座酶的同源性很差,只有C端有一定的相似性。同源建模发现了CasX 蛋白酶C端的RuvC结构域。N端其余的630个氨基酸并没有发现跟其它蛋白有相似性。说明它是一个新的II型Cas蛋白酶,跟 CasX蛋白相连的同源序列有37bp,间隔序列为33-34bp。
Figure3中作者检测了CasX是不是跟Cas9和Cpf1一样具有切割活性,构建了一个含有CasX ,短的间隔重复序列的质粒,然后将它转化到大肠杆菌中,这样一个人工的微型CRISPR-CasX载体能够切割抗性基因,使得大肠杆菌不能在抗性平板上生长(Fig.3a)。 也能靶向切割大肠杆菌增殖调控的基因,影响大肠杆菌繁殖(Fig.3b,3c)。 通过在构建了的含有CasX ,短的间隔重复序列的质粒上PAM序列两端引入随机碱基,分析后发现发现在5’端更偏好为TTCN,3’端没有偏好性。因此(Fig.3d),接着在环境微生物Deltaproteobacteria中发现CasX相连的PAM序列5’端序列是TTCA。进一步验证了上面大肠杆菌中分析得到的结论。
作者又通过大规模转录组分析分析了CasX需不需要tracrRNA靶向切割,发现了非编码的RNA转录本与CRISPR重复序列互补靶向CasX切割(Fig.3e)。又通过Northern杂交寻找了表达CasX 大肠杆菌的非编码RNA序列。发现在跟CasX同一条链110bp长的RNA转录本为介导CasX切割向导RNA。转录组学分析表明 CrRNA最后剪切为23bp重复和20bp的间隔序列。这与 CRISPR-Cas9系统类似(Fig.3f) 为了证明 CasX系统需要 tracrRNA,作者删除前面构建CRISPR-CasX系统中tracrRNA 序列,发现细菌能够在抗性板上存活。(Fig.3g)tracrRNA和crRNA形成sgRNA, 表达 crRNA和短的 sgRNA并不能让 CasX系统起作用,不能切割抗性基因,大肠杆菌能在平板上生长。这些结果说明 CRISPR-CasX系统需要crRNA和tracrRNA形成二聚体靶向切割DNA。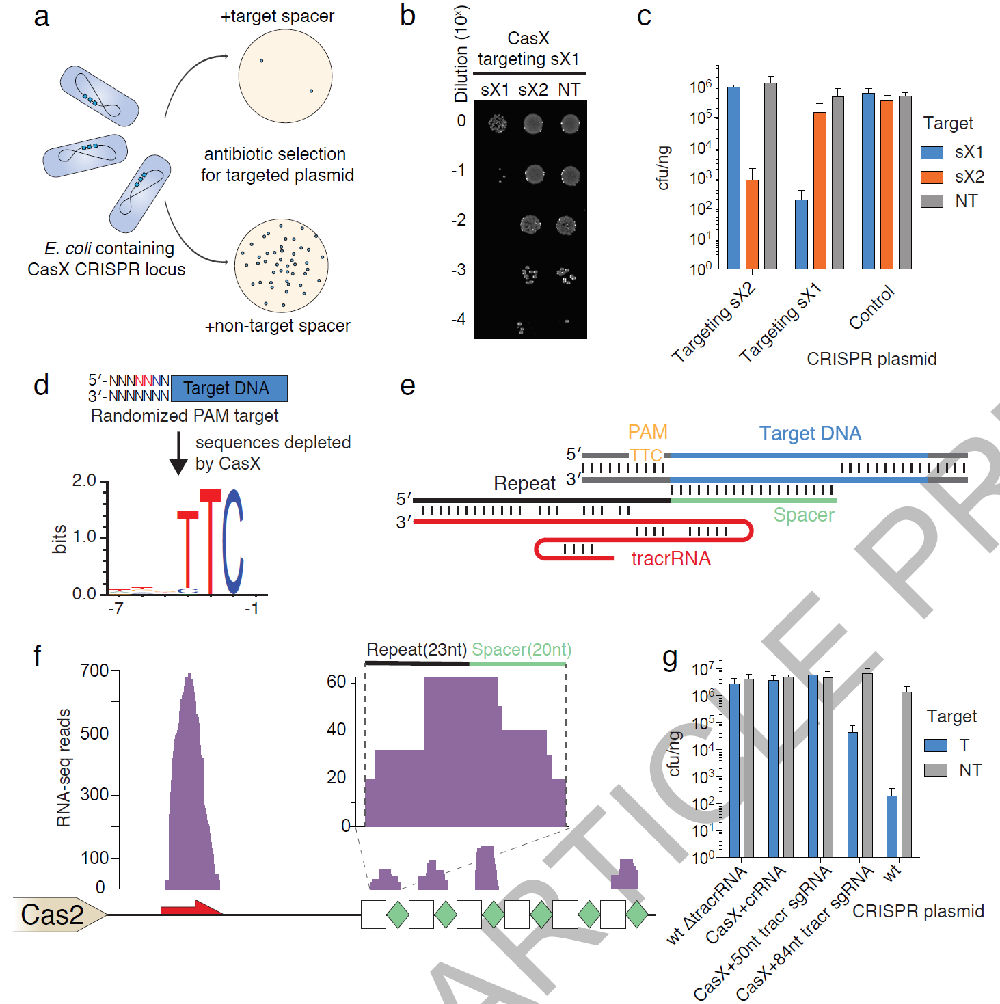
作者在特定种的candidate phyla radiation (CPR)细菌中发现了另外一种 Cas蛋白酶,大约1200个氨基酸序列,命名为CasY,在CRISPR-CasY位点还包括Cas1和CRISPR簇(Fig.4a)。一般的CRISPR间隔序列为17-19bp, CasY.5的间隔序列为27-29bp,根据与重复序列的互补性,作者预测了CRISPR-CasY系统的tracrRNA序列。
6种CasY蛋白在共同数据库并没有检索到同源序列。HMMs建模分析发现6种CasY蛋白中的4种与与C2c3蛋白C末端存在同源性,N末端45个氨基酸存在同源性。CasY跟已有的Cas系统没有很好的同源性,作者想要证明它有没有RNA向导的DNA切割活性。由于它的间隔序列比较短,还没有足够证据说明它需要PAM序列。将微型CRISPR-CasY.1位点人工插入质粒中,转化入大肠杆菌,靶向一个载体含有跟CRISPR簇相应的间隔序列在5’和3’ 端引入随机碱基来寻找可能的PAM序列。发现被切割的序列5’端含有TA。(Fig.4b).根据发现的PAM序列,外源性的启动子驱动CasY.1和另外一个载体含有单个PAM序列, 5’端TA的 PAM序列很容易被CasY切割,说明CasY需要 PAM序列且具有DNA切割活性。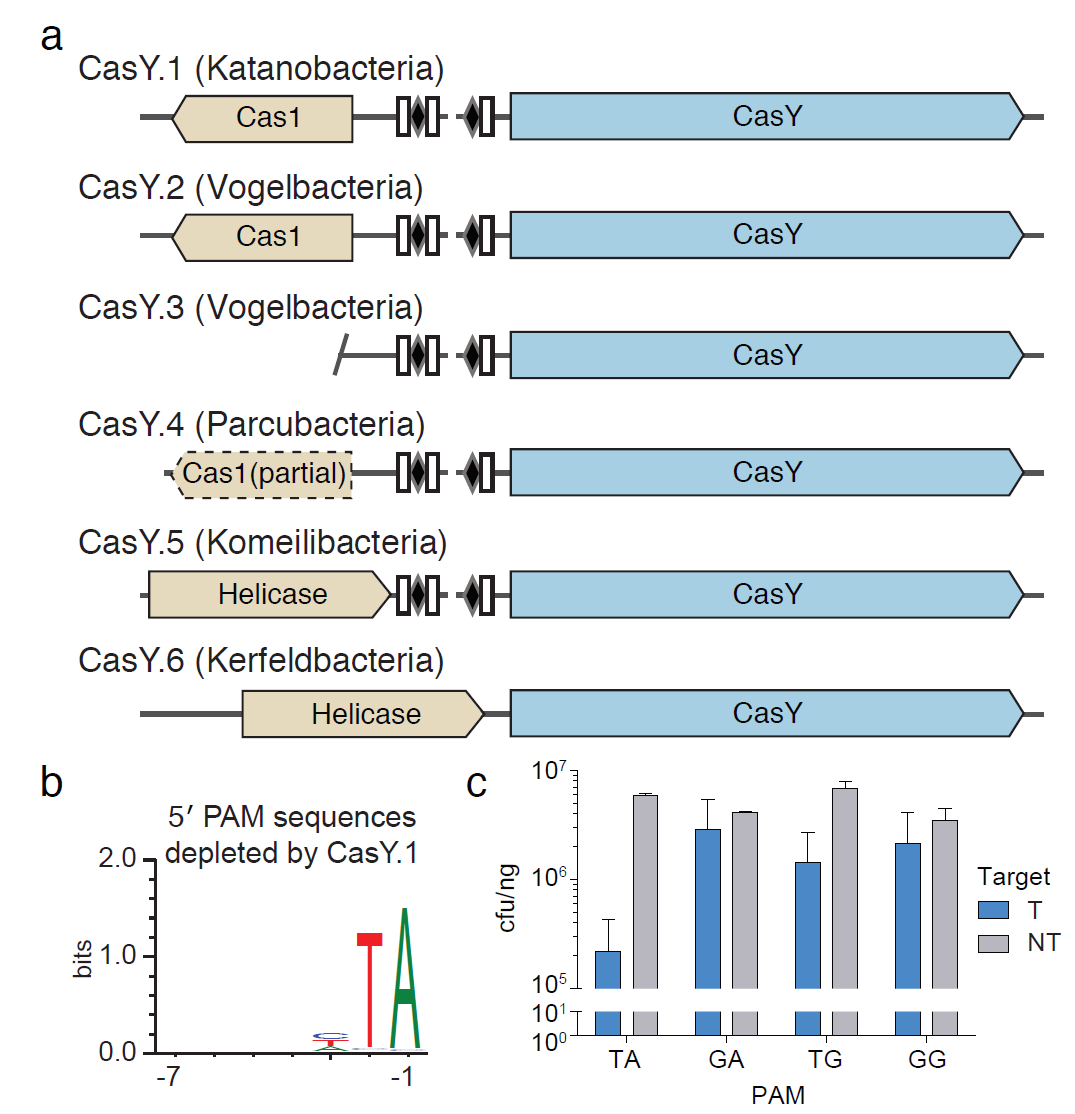
CRISPR-CasX和CRISPR-CasY系统能否成为一个更有效的基因编辑工具,需要广大基因编辑工作者的共同完善和拓展,有CRISPR-Cas9作为模板,相信CasX和CasY的后续工作跟进的会更加迅速。
参考文献:(回复“JC30”下载该文献)
1. Burstein D, Harrington LB, Strutt SC et al. New CRISPR–Cas systems from uncultivated microbes. Nature 2016.
感谢北京物理所颜博士的精彩解读!
我们组建了CRISPR-Cas9专题微信群,欢迎扫码加好友拉入!

还可以微信扫一扫关注我们的微信公众平台:

https://blog.sciencenet.cn/blog-3097940-1022894.html
上一篇:【学术快讯】南通大学刘东Cell Research发文证实NgAgo可用!但是
13 王毅翔 麻庭光 LetPub编辑 黄永义 王天燕 林辉 王志强 孙学军 xchen nuobeier1997 wqhwqh333 liuhaoa1234 htli