ВЉЮФ
ЁОЮФЯзНтЖСЁПNC10 bacteria in marine oxygen minimum zones
||
NC10 bacteria in marine oxygen minimum zones
The ISME Journal 10, 2067ЈC2071(2016)
вЛЁЂеЊвЊ
NC10УХЕФЯИОњЭЈЙ§вЛжжЖРЬиЕФЁЂВњЩњO2ЕФЁЂФкВПКУбѕЕФМзЭщгЊбјаЭЭООЖНЋбсбѕМзЭщбѕЛЏгыбЧЯѕЫсбЮЕФЗДЯѕЛЏзїгУСЊЯЕЦ№РДЁЃдЖбѓжаЕФNC10ЩњЬЌЮЛЩаЮДШЗЖЈЁЃБОбаОПЯдЪОЃЌNC10ЯИОњДцдкгкФЋЮїИчББВПКЭИчЫЙДяРшМгИННќЕФЕЭбѕЧјЃЈOMZsЃЉЃЌВЂЧвзЊТМЛюдОЁЃдкЫљгаеОЮЛЖММьВтЕНNC10ЕФ16S rRNAЛљвђЃЌВЂЧвдкбЧЯѕЫсбЮКЭМзЭщХЈЖШЩ§ИпЕФШБбѕЧјгђжаЃЌЦфЗсЖШДяЕНЗхжЕЁЃМзЭщЕЅМгбѕУИПХСЃЛљвђЃЈmethane monooxygenase genesЃЉЕФЯЕЭГЗЂг§ЗжЮіНјвЛВНжЄЪЕСЫNC10ЕФДцдкЁЃгыNC10ЦЅХфЕФrRNAКЭmRNAзЊТМЮядкOMZФкДяЕНЗхжЕЃЌВЂЧветаЉзЊТМЮяАќРЈЭЦЖЈЕФбЧЯѕЫсбЮвРРЕадФкашбѕЭООЖЕФЛљвђЃЌЦфжаАќКЌNC10ЯИОњвЛбѕЛЏЕЊЃЈNOЃЉЛЙдУИЖРЬиЛљађНсЙЙЕФзЊТМЮяЕФИпЖШБэДяЃЌЫќУЧБЛМйЩшдкВњЩњO2ЕФNOЦчЛЏЗДгІжаВЮМгЁЃетаЉЗЂЯжжЄЪЕСЫдЖбѓOMZsЪЧNC10ЕФЩњЬЌЮЛЃЌБэУїИУзщдкOMZЕЊЁЂМзЭщКЭбѕбЛЗжаЕФзїгУЁЃ
ЖўЁЂжївЊФкШн
ЮЂЩњЮядкМзЭщЃЈCH4ЃЉЕФЯћКФжаЦ№зХЙиМќзїгУЃЌАќРЈгЩЪШМзЭщОњЭъГЩЕФашбѕМзЭщбѕЛЏвдМАгЩвРРЕСђЫсбЮЕФЙХЯИОњЭъГЩЕФбсбѕМзЭщбѕЛЏЃЈAOMЃЉЃЌAOMБЛШЯЮЊЪЧМзЭщбЛЗживЊЕФЛуЃЈReeburghЃЌ2007ЃЉЁЃЕЫЎЩњОГЕФбаОПНјвЛВННЋAOMгыЕЊбѕЛЏКЯЮяЕФЛЙдСЊЯЕЦ№РДЃЈАќРЈЯѕЫсбЮЃЈNO3-ЃЉКЭбЧЯѕЫсбЮЃЈNO2-ЃЉЃЉЃЈRaghoebarsing et al., 2006; Haroon et al., 2013ЃЉ.ЁЃ зюНќЗЂЯжЕФNC10УХжаЛЙдNO2ЕФЯИОњЭЈЙ§ЖРЬиЕФвРРЕNO2ЕФбсбѕМзЭщбѕЛЏЭООЖЃЈn-damoЃЉНЋCH4ЕФбѕЛЏгыN2ЕФВњЩњёюКЯдквЛЦ№ЃЈRaghoebarsing et al., 2006; Ettwig et al., 2009ЃЉЁЃn-damoЭООЖНЋNO2-ЛЙдЮЊNOЃЌетЪЧЕЫЎГСЛ§ЮяжаЕФCandidatus Methylomirabilis oxyferaЃЈбѕЛЏМйЫПНЭФИФюжщОњЃЉЕФЬиЕуЃЈEttwig et al., 2010ЃЉЃЌВњЩњЕФNOБЛМйЖЈЫцКѓЗжНтЮЊN2КЭO2ЦјЬхЃЌЦфжаO2зїЮЊашбѕЪШМзЭщОњФкВПЕФбѕЛЏМСЁЃ
n-damoЕФЩњЬЌбЇвтвхКЭNC10ЯИОњЕФЛЗОГЗжВМЧщПіШдШЛВЛЧхГўЁЃвРРЕNO2-ЛђNO3-ЕФAOMЕФЛЗОГжЄОнжївЊРДздЕЫЎГСЛ§ЮяКЭЪЊЕиЃЈР§ШчDeutzmann et al., 2014; Norei and Thamdrup, 2014; Hu et al., 2014ЃЉЃЌЭЌЪБЃЌNC10ЛљвђдкЖрбљЕФШБбѕЕЫЎЩњОГЃЈShen et al., 2015ЃЉвдМАзюНќдкжаЙњФЯКЃИЛКЌNO2-ЕФГСЛ§ЮяжаБЛМьВтЕНЃЈChen et al., 2015ЃЉЁЃШЛЖјЃЌдЖбѓжаNC10ЯИОњКЭn-damoЕФЧБСІШдШЛЮДжЊЁЃ
ЕЭбѕЧјЃЈOMZsЃЉЪЧвРРЕNO2-ЕФn-damoЕФЧБдкЩњЬЌЮЛЁЃдкжївЊгЩЩЯЩ§СїЧ§ЖЏЕФOMZжаЃЌЮЂЩњЮяЕФКєЮќЯћКФ[O2]ЕМжТШБбѕЃЈ<10nMЃЉЃЈTiano et al., 2014ЃЉЃЌАќРЈЖЋШШДјББЬЋЦНбѓЕиЧјЃЈETNPЃЉжаЪРНчЩЯзюДѓЕФOMZЁЃдкетаЉЧјгђжаЃЌЮЂЩњЮяНЋNO3-ЛЙдЮЊNO2-ЕФЙ§ГЬЛсЧ§ЖЏNO2-ЕНДяЮЂФІЖћХЈЖШЃЈThamdrup et al., 2012ЃЉЁЃOMZЕФЬиеїЛЙдкгкCH4ХЈЖШЕФЩ§ИпЃЈNaqvi et al., 2010ЃЉЃЌЕЭбѕЧјвВШЗЪЕДњБэСЫШЋЧђПЊЗХКЃбѓCH4ЕФзюДѓЛ§РлЃЈSansone et al., 2001ЃЉЁЃОЁЙмвбдкOMZЕФДњаЛзщбЇЪ§ОнМЏжаМьВтЕНгыNC10ЯИОњЯрЙиЕФађСаЦЌЖЮЃЈЭЈЙ§BLASTЃЉЃЈР§ШчDalsgaard et al., 2014ЃЉЃЌЕЋШдШЛШБЗІЖддЖбѓЕЭбѕЧјжаNC10ЯИОњЩњЬЌЮЛЕФШЗШЯЁЃ
БОбаОПЪЙгУЕФбљЦЗЗжБ№РДздДгФЋЮїИчББВПНќАЖЕННќКЃЙ§ЖЩДЉЙ§ETNPЕЭбѕЧјЕФ4ИіеОЮЛЃЈ6ЁЂ8ЁЂ10ЁЂ11ЃЉвдМАЮЛгкИчЫЙДяРшМгGolfo DulceЃЈGDЃЉШБбѕКЃАЖХшЕиЕФ1ИіеОЮЛЁЃБОбаОПбщжЄСЫЁАNO2-ИЛМЏЕФOMZГжгаРрMethylomirabilisбљNC10ЯИОњЁБЕФМйЩшЁЃЃЈВЙГфЭМS1ЃЌВЙГфБэS1ЃЉЁЃдкетСНИіЮЛжУЃЌ[O2]ЖМДгБэУцЯТНЕЃЌдкЩюЖШЮЊ70ЈC130 mЪБНЕжСМьВтЯовдЯТЃЈ20 nMЃЉЃЈЭМ1ЃЉЁЃЖдгкШБбѕЕФOMZЃЌ[NO2-]ЫцбѕЦјЕФЯћКФЖјдіМгЃЌдкETNPЕФЩЯВПШБбѕЧјЃЈ120-150 mЃЉДяЕНЯдзХЕФзюДѓжЕЃЈ4-5 ІЬMЃЉЃЌШЛКѓЫцЩюЖШЯТНЕЁЃдкGDжаЃЌШБбѕЧјгђжаЕФNO2-дк100 mДІРлЛ§жС0.7 ІЬMЁЃДЫДІЕФбЧЯѕЫсбЮдкЕзВПИННќЫЎВу140 mДІЯћЪЇЃЌЦфжаСђЛЏЧтКЭяЇЕФЛ§РлСПдк190 mДІЗжБ№ДяЕН6.6КЭ3.4 ІЬMЁЃ
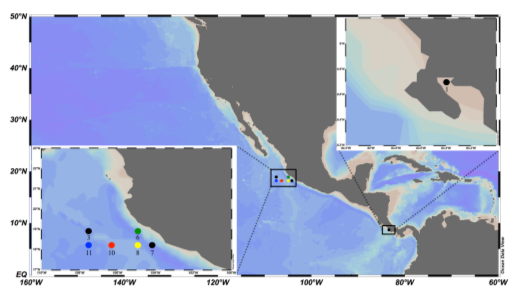
ВЙГфЭМS1 ЗжБ№ЮЛгкФЋЮїИчКЭИчЫЙДяРшМгИННќЕФETNPКЭGDЕЭбѕЧјжаЕФВЩбљЕуЁЃ2014Фъ5дТЃЌЗжБ№дк6ЁЂ8ЁЂ10КЭ11еОЪеМЏСЫгУгкМзЭщХЈЖШЗжЮівдМАn-damoЯИОњЩњЛЏКЭЗжзгЗжЮіЕФETNPбљЦЗЁЃ2013Фъ7дТЃЌдк6КЭ10еОЪеМЏСЫгУгкЗжзгЗжЮіЕФЦфЫћбљЦЗЁЃЭМжаЛЙЯдЪОСЫ3КХеОКЭ7КХеОЃЈКкШІЃЉЃЌЫќУЧдк2014ФъВтСПСЫбсбѕАБбѕЛЏКЭЗДЯѕЛЏзїгУЫйТЪЃЈШче§ЮФЫљЪіЃЉЁЃGDЕФЫљгабљЦЗОљдкGolfo DulceЕФзюББЖЫЕФ1КХеОВЩМЏЁЃ
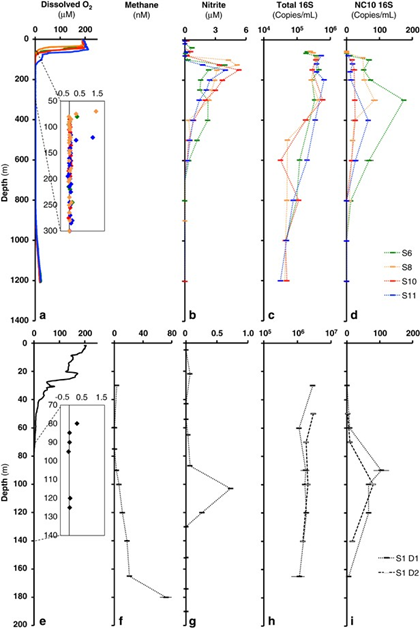
ЭМ1 ETNPЕЭбѕЧј2014Фъ5дТЃЈa-dЃЉКЭGDЕЭбѕЧј2015Фъ1дТЃЈe-iЃЉЕФЫЎЬхЛЏбЇКЭЮЂЩњЮяЩњЮяСПЁЃETNPеОЕФЬнЖШБфЛЏЃЌбеЩЋЖдгІВЙГфЭМ1ДгНќАЖЃЈS6ЃЉЕННќКЃЃЈS11ЃЉЁЃЃЈaЃЌeЃЉШмНтбѕЃЌЛљгкБъзМКЃбѓClarkаЭO2ЕчМЋЕФШмНтбѕЃЌаЁВхЭМЯдЪОЕФЮЊИќИпЗжБцТЪЕФSTOXДЋИаЦїВтСПжЕЃЛЃЈfЃЉМзЭщЃЛЃЈbЃЌgЃЉбЧЯѕЫсбЮЃЛЛљгкЖд0.2ЈC1.6 ІЬmЩњЮяжЪВПЗжЕФqPCRЗжЮіВњЩњЕФЃЈcЃЌhЃЉзмЩњЮяСП16S rRNAЛљвђМЦЪ§КЭЃЈdЃЌiЃЉNC10Ьивьад16S rRNAЛљвђМЦЪ§ЁЃETNPЕФЪ§ОнЗДгГСЫ2014Фъ5дТжабЎЪЙгУCTD /rosetteЛђpump-profilingЯЕЭГЕФРыЩЂЪеМЏЃЈУПИіЩюЖШЃЉЁЃGDЪ§ОнЗДгГСЫЪЙгУЪжЙЄВПЪ№ЕФNiskinЦПЕФРыЩЂЪеМЏЃЈУПИіЩюЖШЃЉЃЌЦфжаD1РДзд2015Фъ1дТ21ШеЃЌВЂгк1дТ23ШеЃЈD2ЃЉЛёЕУСЫЖюЭтЕФ16SМЦЪ§бљБОЁЃЮѓВюbarЪЧжиИДбљЦЗжЎМфЕФБъзМВюЃЈЖдЭЌвЛ[CH4]КЭ[NO2-]ЕФМЦЫуЗжБ№НјааСЫСНДЮКЭШ§ДЮЕФжиИДЃЛгУгк16SМЦЪ§ЕФqPCRНјааСЫШ§ДЮжиИДЃЉЁЃ[O2]Ъ§ОнРДздУПИіеОЕуЕФДњБэадбљЦЗЁЃаЃзМЗНУцЕФЬєеНзшАСЫETNPжазМШЗЕФ[CH4]ВтСПЃЈМћЮФБОЃЉЁЃ
МзЭщДцдкгкETNPКЭGDЕФШБбѕЧјжаЁЃЗНЗЈКЭаЃзМЗНУцЕФЬєеНЪЙБОбаОПЮоЗЈШЗЖЈENTPбљЦЗжаМзЭщЕФШЗЧаХЈЖШЁЃШЛЖјЃЌдкETNPеОЕуЕЭбѕЧјЩюЖШЕФбљЦЗжаШдШЛПЩвдМьВтЕНМзЭщЃЌетгыЯШЧАЕФбаОПвЛжТЃЌМДби26.8 kg m-3ЕШУмЖШЯпЗжВМЕФETNP CH4зюДѓжЕЗДгГСЫCH4РДздгкбиКЃГСЛ§ЮяжаЕФЦНСївЦЖЏЃЈSansone et al., 2001; Pack et al., 2015ЃЉЁЃGDжаЕФCH4ЧњЯпвВАЕЪОСЫетжжCH4ЕФГСЛ§ЮяРДдДЃЌGDЕФ[CH4]ЕФКЌСПдкбѕдОВувдЩЯNO2-ЕФзюДѓжЕЮЛжУДІгкдМ6 nMЕФКлСПЫЎЦНЃЌЖјдкСђЛЏЕФЕзВуЫЎжаДІгкдМ80 nMЕФЫЎЦНЁЃ
ЪЙгУNC10Ьивьад16S rRNAЃЈ16SЃЉЛљвђв§ЮяНјqPCRМьВтЃЌдкЫљгаOMZЮЛЕуОљМьВтЕНNC10 16SЛљвђЃЌЦфжаETNPЕФзюДѓМЦЪ§дкдМ300-400 mЕФЮЛжУЃЈ25ЈC174 copies mL-1ЃЛеМ16SзмМЦЪ§ЕФ0.02ЈC0.05ЃЅЃЉЃЈЭМ1ЃЉЁЃетжжЗжВМБэУїЃЌГ§СЫШБбѕЬѕМўЭтЃЌETNPжаЕФNC10ЯИОњЛЙгыCH4ЯрЙиСЊЃЌЖјВЛЪЧгыРДздЩЯВуЕФгаЛњЭЈСПЙиСЊЁЃвђЮЊШчЙћгыгаЛњЭЈСПЯрЙиСЊЃЌФЧУДЦфгІдЄМЦдкгаЛњгЊбјЛюадзюИпЕФOMZжаДяЕНМЦЪ§ЗхжЕЃЈ(Babbin et al., 2014ЃЉЁЃдкGDжаЃЌдк90 mДІNO2-КЭCH4ИЛМЏЧјгђЕФЖЅВП16S rRNAМЦЪ§ДяЕНЗхжЕЃЈ105 copies mL-1ЃЛеМзмМЦЪ§ЕФ0.006ЃЅЃЉЁЃдкETNP OMZЩЯЗНКЭЯТЗНвдМАGD OMZЩЯЗНЃЌОљМьВтВЛЕНNC10 16S rRNAЛљвђЛђепNC10 16S rRNAОљНгНќМьВтМЋЯоЃЈ6 copies mL-1ЃЉЁЃ
БрТыПХСЃМзЭщЕЅМгбѕУИЃЈpMMOЃЉЕФpmoAЛљвђПЩвдДпЛЏЪШМзЭщзїгУжаЕФCH4бѕЛЏВНжшЃЌЖдИУЛљвђЕФВтађНсЙћНјвЛВНжЄЪЕСЫNC10ЕФДцдкЁЃгУNC10Ьивьадв§ЮяРЉдіПЩДгЫљгаЩИбЁЕФOMZбљЦЗжаЛиЪеЕНpmoAЛљвђЃЌетаЉЩИбЁЕФOMZбљЦЗЪЧвЛИіДњБэСЫЫљгаеОЮЛЕФзгМЏЃЈВЙГфБэS1ЃЉЁЃЖдПЫТЁађСаЃЈETNPжаЕФ26ИіКЭGDжаЕФ2ИіЃЉНјааЯЕЭГНјЛЏЗжЮіЃЌНвЪОСЫгыNC10ЛЗОГПЫТЁжаЕФpmoAКЭCa. M. oxyferaЃЈЭМ2ЃЉгаЙиЕФ4жжУмЧаЯрЙиЕФЯЕЭГаЭЃЈ>98.5ЃЅЕФЙВЯэАБЛљЫсЭЌвЛадЃЉЁЃетаЉNC10ЛЗОГПЫТЁжаЕФpmoAКЭCa. M. oxyferaађСагыФЯКЃГСЛ§ЮяжаЕФађСаОпгазюИпЕФЯрЫЦадЃЈChen et al., 2015ЃЉЃЈChen et alЁЃЃЌ2015ЃЉЁЃетаЉађСаЕФЛиЪеБэУїЃЌOMZжаЕФNC10ЯИОњПЩФмЛсбѕЛЏCH4ЁЃ
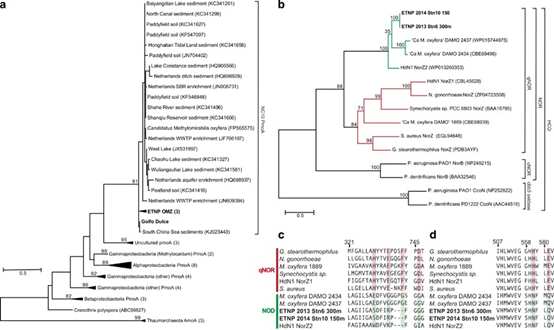
ЭМ2 OMZжаNC10ЕФДцдкКЭзЊТМЛюадЕФЯЕЭГЗЂг§жЄОнЁЃЃЈaЃЉЮЂСЃМзЭщЕЅМгбѕУИбЧЛљAЃЈPmoAЃЉЛљвђЯЕЭГЗЂг§ЁЃДгETNPКЭGDЛиЪеЕФPmoAађСаЃЈn = 53ЃЉдкЦфЫћбаОПЕФНЯДѓNC10 cladeађСаНјЛЏжІжавдДжЬхЭЛГіЯдЪОЃЌЫќУЧгыКУбѕЪШМзЭщОњНјЛЏжІЕФPmoAКЭАБЕЅМгбѕУИЃЈAmoAЃЉЕФЭтШКЗжРыЁЃЪЙгУDayhoffШЁДњФЃаЭИљОн88ИіАБЛљЫсЕФзюДѓЫЦШЛЃЈMLЃЉЗжЮіЭЦЖЯЯЕЭГЗЂг§ЁЃеЙЪОЕФЪЧДѓгк70ЕФBootstrapжЕЃЈздеЙжЕЃЉЃЌвдМАЪ§ОнПтађСаЕФNCBIБрКХЁЃРЈКХжаЕФЪ§зжБэЪОУПИіелЕўНкЕужаАќКЌЕФЮЈвЛађСаЕФЪ§СПЁЊЁЊВЙГфЭМS2жаЯдЪОСЫЫљгаађСаЕФЗжРрЕЅдЊБъЪЖЗћКЭБрКХЁЃБШР§ГпДњБэУП100ИіАБЛљЫс50ИіАБЛљЫсБфЛЏЁЃЃЈbЃЉбЊКьЭбѕЛЏУИЃЈHCOЃЉЕФЯЕЭГЗЂг§зёбEttwigЕШШЫЕФЗНЗЈЃЈ2012ЃЉЁЃЭЦЖЈЕФвЛбѕЛЏЕЊЦчЛЏУИЃЈNODЃЉвдТЬЩЋЯдЪОЃЌАќРЈгЩOMZ NC10зЊТМЦЌЖЮзщзАЖјГЩЕФСНИіШЋГЄађСаЃЈБъМЧЮЊETNPЃЉЁЃгУгкзщзАЕФађСаЪЧЭЈЙ§BLASTXгыNIBI-nrЪ§ОнПтБШЖдЕУЕНЕФгыCa. M. oxyferaОпгазюИпЦЅХфЕФађСаЃЈbit score > 50ЃЉЁЃЛљгкLeКЭGascuel_2008ФЃаЭЃЌЭЈЙ§520ИіАБЛљЫсЕФMLЗжЮіРДЭЦЖЯЯЕЭГЗЂг§ЁЃИУЪїЪЧАДБШР§ЛцжЦЕФЃЌЦфЗжжЇГЄЖШвдУПИіеОЕуЕФЬцЛЛЪ§РДКтСПЁЃЃЈcЃЌdЃЉвРОнEttwigЕШШЫЃЈ2012ЃЉЕФЗНЗЈЃЌqNorНсКЯЃЈcЃЉКЭДпЛЏЃЈdЃЉЖдБНЖўЗгЮЛЕуЕФХХСаеЙЪОСЫгыБъзМqNorЃЈКьЩЋЃЉађСаКЭЭЦЖЈNODЃЈТЬЩЋЃЉађСаЕФБШНЯНсЙћЁЃБрКХжИЪШШШСДЧђОњЃЈG. stearothermophilusЃЉЕФВаЛљЪ§ЁЃвѕгАЭЛГіЯдЪОСЫБъзМqNorађСажаБЃЪиЕФВаЛљЃЌЖјМйЖЈNODађСажавбЗЂЩњСЫАБЛљЫсжУЛЛЁЃ
зЊТМзщНсЙћжЄЪЕЃЌРрNC10бљЯИОњдкOMZФкОпгазЊТМЛюадЁЃЖдРДздШБбѕЩюЖШЕФ5Иі2014ФъETNPбљЦЗКЭ3ИіGDбљЦЗЕФШКТфcDNAНјааСЫФёЧЙВтађЃЌВЂРћгУ2013Фъ6дТРДздETNP еОЮЛ6ЕФ5ИіЯжгаЪ§ОнМЏНјааСЫЗжЮіЃЈЪ§ОнМЏРДздGanesh et al., 2015ЃЉЁЃNC10 16S rRNAзЊТМБОЕФЯрЖдЗсЖШЫцзХбѕЦјЕФМѕЩйЖјдіМгЃЌдкETNPКЫаФЃЈ300 mЃЉКЭGDЕФ90 mДяЕНЗхжЕЃЈВЙГфБэS2КЭS4ЃЉЃЌгызюДѓNC10 16SЛљвђМЦЪ§ЕФЩюЖШвЛжТЁЃдкЫљгаЪ§ОнМЏжаЖММьВтЕНЪЙгУзюЕЭЭЌзцЯШЫуЗЈЗжХфИјNC10ЕФMessenger RNAзЊТМБОЁЃГ§СЫвЛИібљЦЗЃЈ150 mЃЌETNPеОЮЛ10ЃЉЃЌNC10 mRNAзЊТМЮяЕФЯрЖдЗсЖШИїВЛЯрЭЌЃЌгыNC10 rRNAЕФЗжВМЦНааЃЌЫцзХбѕЕФЯТНЕЖјдіМгЃЌВЂдкETNP OMZКЫаФжаДяЕНЗхжЕЃЌеМзмmRNAзЊТМЮяЕФдМ0.1ЃЅЃЈВЙГфБэS2ЃЉЁЃ
OMZЕФРрNC10бљзЊТМЮяПтАќКЌСЫФЧаЉдквьЛЏЕЊзЊЛЏжаОпгадЄВтзїгУЕФЛљвђЃЈВЙГфЭМS3КЭS4ЃЌВЙГфБэS5КЭS6ЃЉЁЃБрТыЖдБНЖўЗгвРРЕадNOЛЙдУИЃЈqNorЃЉЕФСНИіnorZЛљвђЗсЖШзюИпЃЌЙВЭЌДњБэ51ЃЅзюЕЭЭЌзцЯШЗжХфИјNC10ЕФзЊТМБОЁЃЖдгкСНИібљЦЗЃЈETNPЃЌеОЮЛ6ЃЌ300 mЃЌ2013ФъЃЛеОЮЛ10ЃЌ150 mЃЌ2014ФъЃЉЃЌqNorзЊТМЮяЕФзщзАВњЩњСЫШЋГЄЛљвђађСаЃЌИУађСадкЯЕЭГЩЯгыqNorБфЬхЃЈbootstrapЃК100ЃЉГЩДиЃЌгЩCa. M. oxyferaЛљвђзщЕФЛљвђзљDAMO_2434КЭ2437БрТыЃЈЭМ2ЃЉЁЃдкCa. M. oxyferaИЛМЏЪБЃЌNO2-ЛЙдЙ§ГЬжаВЛЛсГіЯжN2OЃЈEttwig et al., 2010ЃЉЃЌОнЭЦВтЃЌDAMO_2434КЭ2437БрТыЕФqNorЙІФмгыNODЯрЫЦЃЌНЋСНИіNOЗжзгзЊЛЏЮЊN2КЭO2ЃЌЖјВЛЪЧВњЩњN2OЃЈEttwig et al., 2012ЃЉЁЃЕФШЗЃЌDAMO_2434КЭ2437дкЖдБНЖўЗгНсКЯЮЛЕуКЭДпЛЏЮЛЕуЕФЙиМќВаЛљДІЖМгаШЁДњЁЃДЫДІЛиЪеЕФqNorађСавВШБЗІБъзМЖдБНЖўЗгНсКЯЛљађЕФБЃЪиадЃЌВЂЧвОпгагыDAMO_2434КЭ2437ЯрЭЌЕФДпЛЏЮЛЕуШЁДњЃЈЭМ2ЃЉЁЃЖдетаЉКђбЁNODЕФМјЖЈдіМгСЫЁАOMZжагабЧЯѕЫсбЮвРРЕадЕФO2ВњЩњЁБЕФПЩФмадЁЃ
дкгЩзюЕЭЭЌзцЯШЫуЗЈШЗЖЈЕФNC10зЊТМЮяжаУЛгаМьВтЕНCH4бѕЛЏЛљвђЃЌАќРЈФЧаЉБрТыpMMOУИЕФЛљвђЁЃЕЋЪЧЃЌгыCa. M. oxyfera pmoBОпгазюИпBLASTXЦЅХфЖШЕФETNP OMZзЊТМБОЃЌдкETNPжаБЛМьГіОпгаНЯЕЭЕФКЌСПЃЌЕЋдкGDжаЮДБЛМьГіЃЈВЙГфБэS6ЃЉЁЃОЁЙмИУЧјгђДІгкбсбѕЬѕМўЃЌдкETNPжаШдЮДМьВтЕНгабѕМзЛљгЊбјаЭЕФpMMOБрТызЊТМБОЃЌЕЋдкGD OMZЕФЩЯВПЃЈ90ЁЂ100 mЃЉжаЛиЪеЕНИУзЊТМБОЁЃCH4бѕЛЏЕФNC10ЗжХфЕФзЊТМЮяЫЎЦНЕЭПЩФмЪЧгЩгкВтађЩюЖШЕЭЛђађСаЗжРрДэЮѓЁЃЛђепЃЌOMZ NC10ЯИОњПЩвдРћгУCH4вдЭтЕФФмСПЕзЮяЁЃ
ЮЊСЫбАевn-damoЕФЩњЮяЕиЧђЛЏбЇжЄОнЃЌБОбаОПЖдOMZЕФЫЎНјааСЫЮЊЦк10ЬьЕФ13CH4КЭ15NO2ШБбѕХрбјЃЌвдВтСПбсбѕCH4бѕЛЏКЭN2ВњЩњЕФЫйТЪЁЃдкETNPЕФЫљгаЗжЮіЩюЖШДІвдМАGDЕФ3ИіЗжЮіЩюЖШжаЕФ2ИіЩюЖШЃЈ100ЁЂ120 mЃЉДІЃЌCH4бѕЛЏЫйТЪОљЕЭгкМьВтМЋЯоЃЈ0.6 nM d-1ЃЉЃЈВЙГфБэS7ЃЉЃЌетгывбЗЂБэЕФЮФеТжаETNPЕФЕЭЫйТЪНсЙћвЛжТЃЌвВгыETNPКЭGDеОЮЛЕФNC10ЛљвђМЦЪ§ЖМЕЭЕФНсЙћвЛжТЁЃЯрЗДЃЌдкGDЕФ90 mДІЃЌCH4ЕФбѕЛЏЕФВтСПжЕЮЊ2.6ЁР0.7 nM d-1ЃЌВЂЭЈЙ§ЬэМгpMMOвжжЦМСввШВБЛвжжЦЃЌетвЛНсЙћБэУїЃЌOMZжаЕФCH4БЛpMMOДпЛЏЕФAOMЙ§ГЬЯћКФЁЃЕЋЪЧЃЌдкДЫЩюЖШДІЃЌДгМйЖЈЕФашбѕМзЛљгЊбјаЭОњжаЛиЪеЕНMMOзЊТМБОБэУїЃЌгЩNC10ВњЩњЕФМзЭщгЊбјаЭЛюадКмПЩФмЗЧГЃаЁЁЃN2ЕФЩњВњТЪДѓДѓГЌЙ§СЫGDжаЕФCH4бѕЛЏТЪЃЌетНјвЛВНБэУїNC10ЖдЙЬЖЈЕЊЫ№ЪЇЕФЙБЯзКмаЁЃЈВЙГфБэS7ЃЉЁЃ
змЕФРДЫЕЃЌетаЉНсЙћШЗЖЈСЫДѓбѓOMZЪЧNC10ЯИОњЕФЩњЬЌЮЛЁЃдкETNP OMZКЫаФДІЕФЫЎЭЃСєЪБМфЙРМЦЮЊ3.9ЁР0.8ФъЃЈDeVries et al., 2012ЃЉЁЃгаСЫетИіЙРМЦжЕЃЌВЂЙлВьЕНGDжаNC10 16SЛљвђЗсЖШЕФЯТНЕЕНгыГСЛ§Юя-ЫЎНчУцЬљНќЕФОрРыЃЌБэУїетРяМьВтЕНЕФDNAКЭRNAађСаВЛЬЋПЩФмДњБэДгГСЛ§ЮяжаЩјШыЯЕЭГЕФвьдДЯИАћЁЃЕБШЛЃЌn-damoЕФЙиМќеяЖЯУИЕФзЊТМЃЌЬиБ№ЪЧгЩqNorБрТыЕФМйЖЈNODЕФзЊТМЃЌБэУїNC10ЯИОњдкOMZЕЊКЭбѕЦјЕФзЊЛЏжаЗЂЛгзїгУЁЃОЁЙмДЋЭГЕФЗДЯѕЛЏзїгУКЭбсбѕАБбѕЛЏзїгУЮовЩЪЧOMZжажївЊЕФЕЊЛуЃЌЕЋгЩNC10ВњЩњЕФN2ДњБэСЫвЛжжИНМгЕФЕЊЫ№ЪЇЭООЖЁЃNC10 pmoAЛљвђЕФЛиЪеЁЂгыCH4КЭNO2-ЯрЙиЕФNC10ЛљвђКЭзЊТМБОЕФДЙжБЗжВМдіМгСЫOMZжаNC10ЯћКФCH4ЕФПЩФмадЃЌОЁЙметвЛдЄВтШдгаД§бщжЄЁЃМјгкдкOMZжаЙлВьЕНCH4ЕФИЛМЏЃЌетаЉИЛМЏРДздГСЛ§ЮяЕФЖдСїЃЈPack et al., 2015ЃЉЛђПЩФмРДздOMZВњМзЭщЮяжЪЃЈВЙГфаХЯЂЃЌВЙГфБэS8ЃЌВЙГфЭМS5ЃЉЃЌМДЪЙOMZ NC10ЯИОњЯћКФCH4ЕФЫйТЪЕЭЃЌвВПЩФмдкПЊЗХКЃбѓЕФМзЭщдЄЫужаЗЂЛгживЊзїгУЁЃ
ВЙГфЭМЃК
ВЙГфБэЃК
41396_2016_BFismej2015262_MOESM84_ESM.xlsx
https://blog.sciencenet.cn/blog-3444937-1248446.html
ЯТвЛЦЊЃКЁОЮФЯзНтЖСЁПЖЋШШДјББЬЋЦНбѓДѓбѓзюаЁКЌбѕДјЩњЮяЙЬЕЊ