博文
传粉者分布、生态功能以及保护措施的多样性
|||
传粉者分布、生态功能以及保护措施的多样性
原文:http://www.annualreviews.org/doi/abs/10.1146/annurev-ecolsys-110316-022919
题目:Pollinator Diversity: Distribution, Ecological Function,and Conservation
作者:Jeff Ollerton, Faculty of Arts, Scienceand Technology, The University of Northampton
翻译:张丹
摘要:传粉者具有重要的生态学功能,在植物的多样性,繁衍生息,与其他生物的关系以及全球农业等方面都非常重要。因此,传粉者无论是在全球生态系统还是食物安全方面都扮演非常重要的角色,而这在生物群体中是独一无二的。在过去的20年,人们对传粉者以及传粉生态的兴趣空前上升,这种现象可能源于人们对传粉者物种多样性和丰富度下降的担忧。这篇综述主要包括传粉者的物种多样性;目前以及更久远以前的分布;其多样性对生态系统功能的作用(包括agro-ecology);当前时间尺度上多样性和丰富度的改变,包括non-native物种;以及传粉者多样性保护的讨论等几个方面。
关键词:蜜蜂,多样性,生物地理学,互利共生,传粉者
1.前言:
传粉者无论是在科学领域还是在人们普遍的意识中都有很重要的位置。一些“拯救蜜蜂”的宣传活动以及植物与传粉者之间有趣的相互作用(在生态学和农业中都非常重要同时也涉及到进化的主要问题)在证明传粉者的重要性方面起了很大的作用。在陆地生态系统中,这是最重要的生态相互作用之一,这种相互作用为大多数依靠传播种子繁殖后代的植物以及推动进化水平多样化的植物和传粉者提供服务。植物本身是静止的,无法主动寻找配偶。因此它们必须通过自花授粉(有近亲繁殖以及遗传多样性丢失的风险的繁殖策略)或者依靠外界的帮助传粉产生后代。这些外界的帮助包括风,水以及动物(包括脊椎动物和无脊椎动物)。在这篇文章中关注的主要是动物传粉。但在一些植物中发现了复杂的传粉系统,有风、动物、称为风虫媒(ambophily)。这也更加确信一些没有记载的传粉方法应该更值得被关注。
大多数的有花植物都依靠动物传粉。最近的一项全球评估显示,87.5%的被子植物依靠无脊椎和脊椎动物传粉,而部分重要的裸子植物也用同样的方法传粉。然而,传粉者的重要性以及多样性也存在误传和偏见,蜜蜂的广泛关注与它们的重要性以及保护需求不成比例。因此,这篇文章的主要叙述传粉者的物种多样性;这种多样性是如何进化;传粉者的全球分布以及为什么这种多样性的保护至关重要。
2.传粉者的多样性
有花植物(裸子植物和被子植物)出现的最早时期大概是侏罗纪中期,约1.7亿年前,是进化生物学的历史标志,在那个时期,有花植物中原始的裸子植物依靠风传粉的方式可能会被复杂的动物传粉替代,因此也出现了被子植物的优势地位和多样性。这是教科书和纪录片中多次出现甚至在最近也出现的一个说法。问题是这个说法是不正确的:最早的被子植物在复杂的环境中进化:昆虫为裸子植物传粉而且对于有花植物昆虫传粉可能是最原始的传粉方式。早期裸子植物的传粉方式在1970s的时候就开始讨论研究。但直到最近昆虫授粉在被子植物中真正的多样性和重要性才开始显现。来自中国,西班牙、俄罗斯的化石昆虫在解释昆虫的口器和传粉的联系,揭示了古代昆虫已经成为传粉者:包括中生代的蓟马,苍蝇,草蛉,蝎蛉以及甲虫。尤其是草蛉和蝎蛉与蜜蜂,蝴蝶,蛾类相比不再是重要的传粉者。
有花植物从白垩纪中期(大约1亿年前)开始的优势地位,似乎与传粉者的多样性有关。例如,2013年,Cardinal和Danforth猜测现存蜜蜂的主要进化分支也始于白垩纪中期到后期。这个预测以蜜蜂主要类群分子系统学为基础。
琥珀和分子系统学的知识也给植物和传粉者之间的相互作用关系提供了更新颖的观点,例如,兰花与其蜜蜂之间的关系至少有1500万年之久,而且兰花大概起源于8000万年前。同样,最近在多明尼加发现了马利筋(夹竹桃科:夹竹桃亚科)的花与其传粉者白蚁的琥珀,这种现象在古生物学中第一次发现,同时也是传粉者和植物相互作用的新发现。毫无疑问,化石记录在未来还会给我们带来更多意想不的惊喜。
2.1目前传粉者的多样性
目前,我们无法准确估计多细胞生物的多样性,已经公布的结果显示,物种的数量大概是从200万到1亿不等,但是较低的估计会更精确一点。其中昆虫是主要物种,也是主要的传粉者;因此对现如今传粉者多样性的估计还是尚未解决的。然而,我们对其他生物类群多样性的估计相对比较准确(尤其是鸟,哺乳动物以及一些蜜蜂),对整体的系统多样性研究和所涉及到的研究物种数量都在逐步增加。表一中可以看到对已记录的传粉者多样性的估计,以及一些研究比较多的类群。Wardhaugh于2015年发表了最新的关于节肢动物作为访花者(而不仅仅是传粉者本身)的综述,但是他的分类单元中包含了捕食性的蟹蛛和螳螂,而这两个物种不是常见的传粉者,因此我在表一中没有列出这两个物种。

多样性最高的传粉类群是鳞翅目,该类群中超过14万的物种都有访花习性,其90%以上的成虫都有功能性口器。鳞翅目的多样性大约是第二大传粉类群膜翅目和鞘翅目的两倍,双翅目是传粉昆虫主要四个目中多样性最小的一个,但在未来随着更多工作的深入以及蝇类真正多样性的发现这个现状可能被改变。剩下其他的类群虽然在某些特定的地理环境条件下对植物有重要的生态作用,但它们整体的多样性都较很低。这个表一将来肯定会发生变化,因为大多数的昆虫还没有被发现描述。例如,Kristensen等2007年的研究显示,鳞翅目的现存种大约为50万,而且大多数为飞蛾,有虹吸式口器。这也引出了一个问题,在文献记载中飞蛾为什么不是最普遍的传粉者,最可能的原因是对飞蛾的研究太少。如果对飞蛾这个类群的研究得到重视,将会发现更多不多传粉者(Devoto et al. 2011, Haber & Frankie 1989)。
脊椎动物中传粉者的多样性也相对较低。这其中鸟类的多样性最高,物种数超过了1000种。夜行鼠也似乎比我们想象的更加重要,尤其是在热带雨林中,但问题是对这个类群的研究程度不及鸟类甚至都不及蝙蝠。蜥蜴也是如此,尤其在海岛上,这里植物和传粉者之间协同进化比较好,而且植物与传粉者之间的相互作用也和陆地上的有很大不同。虽然Ollerton认为,在很久以前鱼可能也有传粉作用,但到目前为止还没有观察到这一现象,Tussenbroek等人2016年对海草传粉的研究可能会让这个现象存在的可能性增加。
总的来说,我们现在预计,大约有35万种传粉者为将近35.2万种有花植物传粉,这是一个很有意思的对应现象(仅仅是一个巧合)。有花植物的物种数量大约为40万,由此可见表一中对传粉昆虫多样性的估计还是太保守,如果Costello估计的真核生物的多样性为500万是准确的,同时考虑到这些物种中至少有100万是真菌,藻类,植物以及原生生物,而且在余下的生物中有很大的一部分是水生生物,那么陆地生物中每10个种中至少有一个物种是传粉者。
所有的这些动物都可能是有效的传粉者至少对于它们访问的植物来说是这样的。但不同的授粉者授粉的有效性也有区别,这种不同主要表现在三个方面:群体中动物丰富度的差异;动物对花粉偏好性的不同;动物是否会落到相同物种的花上以及它们移动距离的不同(Herrera 1987,Rodriguez et al. 2013)。造成这些差异的最主要原因是传粉者数量的预计;其次是传粉者的质量。虽然对传粉者重要性的预测是从多样性较低的生物开始,但是这种预测很耗费时间而且很难做到整个群落水平,全球水平上的预计就更不用说了(Ballantyne et al. 2015)。此外,种内传粉能力的差异也让这个问题更加的复杂,例如,个体较大的雄峰相对于个体较小的来说,每次访花时携带的花粉更多,而且每天的活动时间也较长(Willmer & Finlayson 2014)。
另外一种获得不同传粉者相对重要性的方法是从植物的角度出发。图一展示了在32个植物群落系统中,不同传粉系统的平均相对频率(see Supplemental Materials)。这些数据包含了北极,温带,亚热带以及热带的植物系统。毫无疑问的是,较高的标准差和平均值为代表,可以看出区域间存在生物地理差异,但在一个小的数据集(代表了大多数当前可用的数据)中我们法法解决去这个问题。结合图一我们可以看到,最主要的传粉类群是蜜蜂,其次是苍蝇。其他的昆虫在传粉中的重要性相对较低,但毫无疑问它们对植物的传粉也有重要的作用。蝴蝶和飞蛾的重要性不是很明显,这可能是因为对蛾子的研究还不够。图一的结果与表一的物种丰富度没有关系,但也告诉我们,不同类群作为传粉者传粉有效性的差别:蜜蜂和苍蝇的多样性远低于鳞翅目,但是它们浓密的体毛让它们成为有效传粉者传粉者。另外,蜜蜂是唯一一个成虫和幼虫都依赖于花资源的类群。
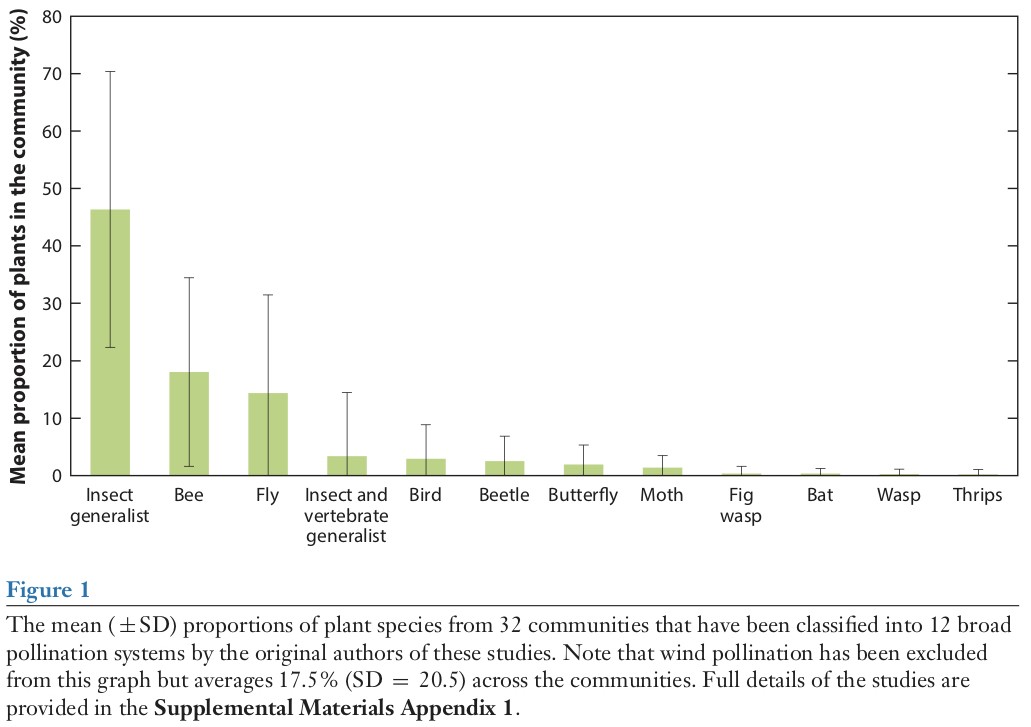
生物多样性名录是全球对生态系统保护结果的重要部分,而且长期以来我们已经意识到,在实施保护的措施之前理解生态系统的功能很关键。植物—传粉者之间相互作用的关系从保护生物学角度来说有两个方面,第一,传粉者的多样性植物繁衍方面非常重要;第二,长期以来传粉者的多样性在陆地生态系统中的作用。文章前面的部分给人感觉是在说动物作为传粉者的多样性以及大部分的植物都需要访花者帮助它们传播花粉,因此,在第四部分我们探讨了这种多样性的重要性以及如果这种多样性丧失对整个生态系统的影响。但目前植物和它们的传粉者之间的相互作用关系还没有被完全发掘,研究者继续发现新的传粉系统的比例将不会下降。2016年甚至发现了一个被证实地传粉系统,包括海草,访花的甲壳类动物以及多毛虫等都在描述之列(Tussenbroek et al. 2016)。
生物多样性的研究可能会忽略物种之间相互作用的多样性,但目前我对物种之间相互作用多样性的研究已经有很长的历史了。尤其是植物—传粉者之间的相互作用,自18世纪以后,一些研究者们就已经开始关注这个问题,其中包括一些很重要的科学家,例如 Joseph Gottlieb Koelreuter、Christian Konrad Sprengel、Charles Darwin和Hermann Muller (Waser 2006)。一个重要的问题是,在这个时期以后,是否全面探究了植物—传粉者之间相互作用多样性,或者说我们现在是否仍然处于发现和分类的阶段。全球分类学中物种积累的增长对于某些物种已经趋于平稳,这并不适用于所有的物种(Costello et al. 2013)。尽管植物与传粉者相互作用的研究历史已经有200年了,但是我们对这些相互作用的总体模式的理解,以及包含的物种的探索还有待完善。
传统意识上对于传粉者的预测都以蜜蜂、蝴蝶以及食蚜蝇作为主要传粉者。事实上传粉系统包括一些不常见或者意想不到的类群,例如:花金龟,捕食蜘蛛,胡峰,蜥蜴,啮齿类动物等依旧认为是例外,但实际上一些认为不可能的种也可能成为主导(Johnson 2004,Olesen & Valido 2003,Ollerton et al. 2003,Shuttleworth & Johnson 2009),而且传粉者类群的范围可能比我们现在认识到的还要广。同时这也引出了一个问题:植物——传粉者之间的相互关系在热带和高纬度地区有什么不同,这是在过去几年中越来越引起人们兴趣的一个问题,在第三部我也探讨了这个问题。
3.在空间领域传粉者的多样性
在空间领域,传粉者多样性是不确定的:无论是对于来自世界不同区域传粉者的数量,还是它们的分类学地位,都具有重要的生物地理结构。特别是传粉者生物多样性会随着纬度和陆地的变化而变化,这反映了地球不同地区的多元化演化历史。但是,目前一些主要的地理差异已经影响了我们对不同传粉群体的相对重要性的理解,而且还存在很多研究没有涉及到的区域。同样,对很多引入传粉者带来的潜在影响我们还知之甚少。
3.1 传粉者的多样性和分布的生物地理学模式
一般来说,传粉者生物多样性遵从预期的物种丰富度随着纬度增加而增加的模式:正如我们所料,由于热带地区具有较丰富的植物群体,所以热带地区的传粉者也更胜一筹(排除一些例外),这二者之间是相互关联的(见Section 6.2)。然而,人们早就蜜蜂作为传粉者中的一个主要群体,热带地区并不是其多样性最高的地区,而是在干燥的亚热带的地中海类型的区域(见图2)(Michener 2007,Ollerton et al. 2006)。需进一步预计更完整的热带地区蜜蜂多样性列表以便证实这种模式(Michener 2007)。相反地,其它传粉者类群(非蜜蜂)却很符合这种普遍发生在热带地区的最大生物多样性模式;例如印度-澳大利亚动物群与古北区的类群相比,新热带地区的鳞翅目物种数目几乎是新北区鳞翅目的五倍(单位面积的物种数量),尽管非洲鳞翅目昆虫多样性几乎是古北区的两倍之多(Kristensen et al. 2007)。
全球某些区域例如南非(Johnson2004),蜜蜂的多样性显著低于预期值。这似乎是植物寻找特殊(从北温带的角度来说)传粉者的演化结果,这些传粉者例如甲虫、黄蜂以及啮齿动物群体,而这些动物在世界的其他地方几乎不作为专门的传粉者存在(Johnson 2004; Ollerton et al. 2003,2006; Shuttleworth & Johnson 2009)。同样,在澳大利亚植物已经与通常意义上的非专业传粉者之间形成了新的传粉关系(Armstrong 1979)。在北极,苍蝇是主要的传粉者(Tiusanen et al. 2016),在北半球除北极以外的大部分地区以及南美,熊蜂属Bombus物种是更具优势的传粉者。而在撒哈拉以南的非洲地区,即使有的物种已经成杰出的传粉者,但某些属的物种仍然欠缺,这依旧是未解难题(见Section 3.4)。
另一个著名的传粉者生物地理分布模式是,在大部分大部分欧洲地区以及地中海区域(少许例外),鸟类是本土植物的传粉者,这是一种比较少见的现象,而世界的其他地区包括不同访花鸟物种(例如美国蜂鸟,非洲、亚洲和澳大利亚的太阳鸟,还有澳大利亚的食蜜鸟;Cronk & Ojeda 2008)。在欧洲,专性传粉鸟类相对匮乏,而在渐新世期间蜂鸟存在的历史原因尚不明朗。然而,许多非专性授粉的雀形目物种,总是携带多种多样的本土植物和外来引进植物的花粉(da Silva et al. 2014),所以鸟类传粉可能更加普遍,只是目前我们还没有认识到或者还没有相关的记录而已。
3.2 在热带地区,植物与传粉者之间的相互作用有何不同?
从两极到热带物种丰富度有增加的趋势,这在18世纪就已经成为共识(Hillebrand2004)。而更高的物种丰富度则表明,热带地区物种之间的相互作用过程,可能会或由于资源竞争(包括相互影响的物种)而使得专一化的比例更高,这是在狭窄的生态幅中不断演化的结果(but see Moles & Ollerton 2016,Vazquez & Stevens 2004)。平均地,在热带地区,专性植物-传粉者相互作用的功能多样性更高(Ollertonet al. 2006),这可能是由于热带地区蜜蜂多样性相对较低(图2;见我在Section3.1关于亚热带南非的评述)。然而,对热带地区的植物-传粉者相互作用更加专一化的这个假设的检验很少,而这些检验都有不同的结果(e.g., Olesen & Jordano 2002,Ollerton & Cranmer 2002)。近期,Schleuning等人(2012)从生态学观点出发,揭示了热带植物与其访花者之间的网络关系实际上缺乏专一化趋势,然而Pauw & Stanway(2015)则拿出了证据,证明热带地区专一化相互作用增加的趋势发生在南半球(而不是北部)。他们认为南半球气候的长期稳定性以维系这种专一的相互作用(see also Dalsgaard et al. 2011)。

以我们目前的知识理解生物传粉,其作为一种生态功能在某些区域可能更容易被破坏,如在北温带和南半球,而不是热带区域作为整体,对干扰的敏感性是一个地理上的错综复杂因素而不是仅从纬度就可以预测的(Vizentin-Bugoni et al. 2017)。
3.3 非本地传粉者的引入
某些地方通过引入非本土物种特别是蜜蜂(Goulson2003,Russo 2016),人为的增加传粉昆虫的多样性,有目的地或偶然地提高作物授粉情况。在一些海洋岛屿上,引入传粉者物种的数量要比本土传粉者数量多;例如,一个最近发表的亚速尔群岛Azores蜜蜂名册显示,该岛的19个记录种(包括蜜蜂蜂巢的管理)中只有4种可能是来自本土(Weissmannet al. 2017)。如此高的外来物种比例是异常的,而且引入传粉者的数量在调查结果中也有增加,以至海洋岛屿似乎特别容易被入侵(Olesen et al. 2002)。
这些引入种的生态效应从有害到有益,这都取决于引入物种,引入的环境背景以及(特别是)本土传粉物种的多样性(Russo 2016)。最具有影响力的引入种是西方蜜蜂(Apis mellifera),西方蜜蜂农业授粉上的作用远超出其本土领域。大量的研究表明西方蜜蜂的传粉能力超过本土蜜蜂和其它访花者,尽管这与地貌结构是否单一、均匀或者复杂、多样化,以及关于可供蜜蜂觅食的半自然栖息地的数量等都有关(see Herbertsson et al. 2016)。研究也表明蜜蜂能很好地融入到局部传粉网络中,且没有明显的负面效应,至少在较低密度的情况下是如此(Watts et al. 2016)。
其他的引入蜜蜂包括B. terrestris,在非原产地,该种被认为是一个入侵物种(Dafni et al. 2010),而且它还与本地熊峰的丢失有关(Morales et al. 2013)。保护生物学家关心的不仅仅是竞争的直接效应,还担心引入的蜜蜂带来的疾病可能会在本地物种中传播(e.g.,Arbetmanet al. 2013)。
当然引入传粉者也有积极的一面,大量的研究表明非本土的传粉者会促进一些由于缺乏本地传粉者而受威胁植物的繁殖。例如在二十世纪八十年代,Cox(1983)确定了一种夏威夷攀岩植物(Freycinetia arborea)的传粉者是一种外来引入鸟-暗绿绣眼鸟(Zosterops japonica),伴随着本地鸟类传粉者的灭绝。十年前,Lord(1991)揭示了在新西兰,一种负鼠(Trichosurus vulpecula)作为传粉者取代了两种本地蝙蝠(一种非常罕见,另一种可能已经灭绝)。最近,Fox等人(2013)表明,过去九年对北美受到威胁的西部草原上的研究发现,带穗兰花(Platanthera praeclara)的专性传粉者是一种天蛾(Hyles euphorbiae),这种天蛾最初是为了控制入侵植物而引入到本地的。然而,这种引入蛾数量上可能已经超过了本地兰花的传粉者。
传粉者引入后的结果主要取决于当地的环境,当然这些环境会随着时间的推移而改变,例如当前的良性引入物种在未来可能会出现问题。因此,禁止引入非本土熊蜂到澳洲大陆为温室作物授粉一个明智的决定,为了塔斯马尼亚岛Tasmania免受影响(Hingston2007),而且事实上,这种商业作物完全可以由本地蜜蜂完成传粉工作(Hogendoorn et al. 2006)。
3.4 传粉者扩张和收缩的自然范围
除了一些认为因素对物种分布的影响外,我们也预计了一些自然发生的改变,因为物种的扩张和收缩与资源可利用性、天气、扩散机会等的变化(当然,其中一些也可能是人为介导的因素)有关。这种自然变化在英国新世纪时期就已经出现,在英国,许多没有记录的蜜蜂物种数量已经趋于稳定。最完整的文献记载是关于ivy bee (Colletes hederae)和三种熊峰(Bombus hypnorum)的研究,于2001年开始(Goulson & Williams 2001,Roberts & Vereecken 2010),跟踪过程由BWARS社团负责。2015年Rasmont等人的研究显示,熊峰在英国的扩张恰巧在欧洲广布物种扩张的下一阶段。这不是唯一的例子,Rasmont等人(2015)也发现亚洲熊蜂Bombus schrencki已经扩张到了西方的波兰和芬兰,Lopez-Uribe和Cane(2016)记录了南瓜专性传粉蜜蜂(Peponapis pruinosa)的扩张与作物驯化的平行过程,并且Russo(2016)引用了其他北美的例子。
这样的扩张幅度并不惊讶,因为我们早就知道黄蜂和熊峰蜂后能在开放的水域进行数十公里长距离扩散事件(Mikkola 1984)。相比之下,人们对自然收缩的幅度的认识较少,一部分原因是人类自以为是地认为任何一个物种数量的局部下降都有人为因素的干扰,这显然是不可信的,化石记录已经在时间尺度上告诉我们一个物种进入并贯穿整个区域的时间幅度。但是将自然因素从人类主导的地貌分离开来是不可能的。在十九世纪五十年代,英国的23种蜜蜂和访花黄蜂被告知即将灭绝,几乎可以肯定一个或更多的物种已经消失了(Ollerton et al. 2014)。的确,在修订这个文章的很长一段时间,一个物种被再次发现并且很自然地从大陆再度迁移到英国。以前发现的某个类群的化石记录显示,以前有该物种的地方物种已经不再存在,而且它们祖先生存的地方完全出乎意料。近年来研究表明,蜂鸟这种长期以来只人为在新世界中发现的传粉者,竟然出现在欧洲大约三千万年前的渐新世纪(Louchart et al. 2008,Mayr 2004)。这些发现让我们的未知领域变得更多,同时目前还需要更多传粉者和植物之间相互作用的生物地理和生物多样性的研究。
相比当前传粉者在时间和空间上多样性的预测,在全球,地区以及地域尺度上多样性的下降可能更加重要,更加紧迫,这种变化可能意味着传粉的生态功能和生态系统服务功能的下降。在这部分、第5部分,我将概述这些内容。
4.1 授粉者多样性降低的基本证据
很多重要的研究和综述都概述了授粉者多样性和丰富度的减少,例如,Biesmeijer (2006),Potts (2010a),以及Lebuhnet(2013)。但是,对这些物种估计的准确性,传粉者多样性降低(尤其是蜜蜂)是否是文献记载和媒体夸大叙述等方面有不同的意见(Ghazoul 2005)。2015年Goulson和他的同事给出了导致蜜蜂数量下降的证据,同时人们对他们的研究结果也有质疑,尤其是传粉者多样性下降的证据不足(Ghazoul 2015),作者做了相应的回应(Goulson et al.2015b)。从某些方面来说,这不是一个令人满意的交流,交流的重点是农业传粉者而不是所有植物的传粉者。从更加广泛的层面看待传粉者发现:多样性下降的证据更加明显,传粉者的多样性与丰富度在区域和全球范围内都有所下降。
2015年Ghazoul最突出的声明是“在欧洲和北美,传粉者数量多样性下降的证据几乎都是以蜜蜂和蜂为重点”。这个声明更深层的意思是对全球生物多样性下降的担忧是以分类学和地理学为基础,因此证据的涉及面比较窄。总结目前已有的研究显示,很显然事实不是这样的。虽然这个总结并不全面,但是显示了目前分类学和地理学证据的深度和广度。
4.1.1野生蜜蜂(包括熊蜂,独居型以及社会型蜜蜂)
这些物种的多样性和丰富度已经有所下降,而且在英国和欧洲大陆Biesmeijer et al. 2006,Nieto et al.2014,Ollerton et al. 2014),南美(Morales et al. 2013),亚洲(Williams et al. 2009),南非(Pauw 2007),北美(Burkle et al. 2013,Cameron et al. 2011)已经有文献记载。在美国,相关部门已经将夏威夷的全部7种蜜蜂列为濒危物种,这是蜜蜂第一以这种形式在美国出现。
4.1.2 蜜蜂
北美和欧洲的一些地方都有蜂群数量下降的记载,但在别的地方蜂巢数量保持稳定可能还会增加(see Potts et al. 2010b)。尽管全球蜂巢的数目在增加,但是有证据表明,蜜蜂的授粉依然供不应求(Aizen & Harder 2009)。
4.1.3 食蚜蝇
食蚜蝇多样性的降低在荷兰和英国都已经有文献记载(Biesmeijer et al. 2006).但是其他地方的现状还不是很清楚。
4.1.4蝴蝶和飞蛾
鳞翅目的物种多样性和丰富度在英国已经开始下降(Fox 2013,Thomas et al. 2004),在南美,大约有60个物种已经被列为易危,濒危或极度濒危物种,而且特别值得关注的是帝王蝶。同样,环保人士担心,在世界其他地方蝴蝶也是主要濒危对象(例如南非,澳大利亚,欧洲)。
4.1.5访花胡峰
在英国已经发现多刺黄蜂的物种多样性已经有所下降(Ollerton et al. 2014)。但是其他国家还没有发现,尽管非常重要,但是在其他国家该类群不像其他传粉者那样被广泛关注(Ollerton et al. 2003)。人们现在也开始关注气候变化对榕小蜂的影响(Jevanandam et al. 2013)。
4.1.6鸟类和哺乳动物
2015年Regan借助世界自然保护联盟红色名录估计了全球主要哺乳动物传粉者的物种多样性。作者认为,在过去的几十年里,平均每年有2.5个物种被列入该名录,接近灭绝。已有报道称,岛屿上的授粉鸟类已经出现灭绝,例如,夏威夷和新西兰的两种授粉蝙蝠,其中一种在1960s可能已经灭绝,另外一种的数量也急剧下降。
当然,上面提到的这些研究表明,一些物种相比其他物种更具传粉的优势,而且这些分类群的数量相对稳定甚至有所上升。另外,在欧洲西北部传粉者减少的速度也在下降(Carvalheiro et al. 2013; but see Ollerton et al. 2014)。但总的来说,目前在大范围尺度上,传粉者物种多样性和丰富度的下降还没有确凿的证据。
现在人们已经开始普遍的关注引起这种变化的下原因,但是土地使用方式的转化以及农业集约化以后土地的管理导致动物栖息地的丧失是最有可能的原因(Bartomeus et al. 2013,Goulson et al. 2015a,Ollerton et al. 2014,Potts et al. 2010a)。最近关于气候变化对传粉者多样性变化的预测显示,未来传粉者多样性的下降和分布范围的变化仍然存在。
4.2 气候变化与传粉者多样性
一直以来,访花昆虫都是研究气候变化对物种分布影响的主要内容,1999年Parmesan等人首先从蝴蝶开始这个研究(Parmesan et al. 1999,Settele et al. 2008),最主要的原因是相对于其他类群昆虫可以获得更多的数据,并不是因为它们是传粉者。最近,在北温带地区,对蜜蜂包括熊峰属的研究,可能是对植物传粉者单一属的最重要的研究。这个属一直是“欧洲熊峰气候风险和分布区域”研究的主要对象(Rasmont et al. 2015)。主要表现在熊峰对寒冷气候的适应,尤其是它们浓密的体毛,这也可能是它们成为优秀传粉者的原因。根据这一点,我们假设熊峰对气候变化可能很敏感(虽然其他的蜜蜂可能会从某地的气候气候变得温暖和干燥中获益)。2015年,Rasmont和他的同事,试验气候变化对56种熊峰的影响,结果显示36%的物种面临的气候风险较高,41%的物种正处在危险之中(消失率为50%-80%),略高于5%(三个物种)的物种扩大其在欧洲的分布范围。总的来说,在2050年之前,34-52种熊峰适合生存的气候条件将会下降,而到2100年将会扩大到49-55个种。在欧洲南部,熊峰的分布已经较少,因此这种气候的改变最应当引起关注。到2050年,西班牙和葡萄牙可能只存在一种熊峰,这对水果,野生植物以及栽培植物将产生有很大影响。
在极地地区,人们最初认为人为造成的气候变化对生态有影响。1996-2009年,北极圈的开花季节开始变短,访花者的丰富度也开始下降(Høye et al. 2013),尤其是影响力最大的一个类群(双翅目蝇科),它们是仙女木属的主要传粉者,仙女木属也是这个区域主要的蜜源植物(Tiusanen et al. 2016)。但是,热带的物种可能对热量有严格的要求,因此很容易受到全球变暖的影响(Jevanandam et al. 2013)。
推测授粉者对气候变化的反应,主要依赖于物种对气候变暖的适应性以及在新区域适应和扩散的能力。物种保护计划显示,传粉者应当具备在经过一定恢复的区域间迁移的能力,在很多国家这被认为是传粉策略最有影响的规则,例如英国,美国,爱尔兰。在区域水平上,很多实际的保护监控,概念验证工作正在重建以及预测栖息地的多样性,例如欧洲的石楠(Forup et al.2008),北方的松林(Devoto et al. 2012),垃圾填埋场(Tarrant et al. 2013),城市中心(Baldock et al.2015,Sirohi et al. 2015)以及热带岛屿和孤山(Kaiser-Bunbury et al. 2017)。非政府组织的倡议,例如英国的Buglife’s B-Lines 计划,美国的Xerces Society等都在不断地努力。但是,有的策略在阿尔卑斯山和北极地区是不可行的,这些地区传粉者被可移动的范围限制。
5.为什么保护传粉者的多样性很重要?
关于保护传粉者多样性的争论通常包括三个主要因素即实用性、理论性或者观点的抽象性。首先,多样化的生态和农业功能,避免农作物发育不良,传粉者贡献突出。第二,多元化的类群为将来提供生态保障,万一某个关键传粉者种群下降或者灭绝。最后,物种多样性本身就是人类、以及地球生物文化遗传的一部分。这些不是相互排斥的争论,确实反映了目前的紧张局势,传粉者多样性保护是保护生物学家和生态学家提出的为了社会发展的策略。反过来,在自然和生态系统服务(NC-ES)下也可以提出这三个争论,在过去的二十多年,NC-ES已经得到社会的广泛认可(Maceet al. 2012)。然而,一个未解决的问题是,NC-ES对农业服务的争论并不总是反映生物群落的生态现状,这个生物群落中物种稀有性和专一性是生态复合体的一个重要组成部分。
5.1 专一性-普遍性的连续统一体
在过去的30年,有很多关于植物-传粉者相互作用的专一性-普遍性的连续统一体的研究,都表达了不同的观点(Brosi 2016,Fenster et al. 2004,Jordano 1987,Waser et al. 1996)。最近,这个工作由Armbruster (2016)修订和探讨,Armbruster将其描述成“我们思考的主要问题,不仅仅与植物-传粉者相互作用和传粉者服务的生态有关,而且还包括生殖隔离、物种形成、灭绝和群落的集结”。同时,这也是传粉者为什么必须受到保护的核心内容。
这种连续统一体的一个极端是高度专一化,即植物和它传粉者之间一对一的关系。这种关系包括一些相互依赖的授粉系统,例如无花果和无花果黄蜂、叶下花和叶下花蛾、丝兰和丝兰蛾等之间的关系,这些匹配关系从种子寄生关系演化而来(Hembry & Althoff 2016)。另外当涉及到昆虫不访花的欺骗行为时,它们没有相互依赖,例如吊灯花属Ceropegia spp. (Apocynaceae)与其双翅目传粉者之间的关系(Ollerton et al. 2009)。这种连续统一体的另一个极端是植物的花和所吸引的大范围传粉者之间的普遍性。例如一些北美的乳草(Asclepias)物种因能被100多种不同的昆虫传粉而出名(Ollerton& Liede 1997),例如熊蜂和蜜蜂(Bombusand Apis)的传粉者可以为一个群落中大部分的植物授粉,还有一些岛屿上的传粉范围特别广的传粉者(Olesen et al. 2002)。
对于植物或传粉者分类并将其置于这个连续统一体中,取决于对它们之间相互作用观测的空间和时间尺度。我们希望得到的是广布种在它们目前分布区域的专一性而不是考虑到它们全部分布范围的普遍化(e.g., Gomez et al. 2013,Herrera 1988,Ollerton et al. 2009)。同样地,花和传粉者之间关系的变化,依赖于物种的相对丰度。
从时间和空间方面去理解物种相互关系的如何做到专一性和普遍性,在生态取样方面围绕以下几个问题:如何知道已经用足够的时间去理解一个研究系统?取样不足会带来一个植物或传粉者专一化水平的错误结论,但过多样本的收集几乎是不可能的,除非在相互作用很快达到渐近线(平衡)的情况下,但这需要花费一些时间。
大部分植物和传粉者都处于专一性-普遍性连续体之间的位置,而且相比专一性更接近普遍性,至少在生态特化方面是这样的。这意味着(至少在理论上),在大多数植物和传粉者的生态学中存在一定程度的冗余,就好比特定的传粉者可以从不同类型的花中获取资源,反之一个特定的植物也可以被任何一个访花者授粉。对传粉者多样性局部缺失与这种冗余之间的重要性的研究正在展开,通过传粉者互斥实验,得到了不同的结论(见Section 5.2)。
对植物传粉者多样性的一个很普遍的是:“传粉者”的调查通常是花的访问者而不是真正意义上的传粉者。这种批评尤其针对在植物-访花者集合体网络的研究,而这种批评也有一定的效果(但值得注意的是访花行为而不是植物最终的结果,而从访问者的角度来看通常是一种积极的相互作用)。不幸的是,很少有对某一植物访花者和传粉者之间对应关系的数据可以用来理解哪部分访问者是真实有效的花粉载体。因此,一个重要的问题是,对传粉者的调查经常忽略除蜜蜂以外的其他传粉者(假设“蜜蜂是最重要的授粉者”)(Rader et al. 2015)。
萝藦科植物(Apocynaceae:Asclepiadoideae)可以有效解答这个问题,它们的花粉是以花粉粒黏在动物身上的方式进行传播,这使得有效传粉者的鉴定工作相比于其他植物群体更加容易(Ollerton & Liede 1997)。图3a展示了观察到的(这些花来自于南非的夸祖鲁-纳塔尔省KwaZulu-Natal草原上的8个萝藦科物种)访花昆虫的数量和证实过的传粉者的数量关系。很明显,大部分有一个或者两个访花者的专性植物被所有或者大多数访花者授粉,如果所有的访花者都是传粉者,那这个关系的比例就接近1:1。但随着物种数量逐渐增多更普遍化,出现了访花者中传粉者的比例较低的情况。事实上,对于大多数植物只有不到三分之一的访问者才是有效的传粉者(图3b),尽管这被认为是保守估计,就像有的昆虫没有观察到携带花粉的现象,可能是因为低访问频率或者样本限制。值得注意的是,这些访花者中只有很少一部分是蜜蜂:黄蜂,甲虫以及蝴蝶更为常见(see Ollerton et al. 2003,Shuttleworth Johnson 2009,and comments regarding southern Africa in Section 3.1)。

只有维持物种多样性才可能有物种的冗余:在物种已经开始灭绝的群落中,尤其是在某些物种稀缺的时期,传粉者缺失可能意味着一个植物种群的衰退,或者说缺少了一个至关重要植物会导致一个传粉者种群的衰退。
这种现象在单一作物农业生态系统中最极端的例子中可以看到,即这个农业生态系统失去了本土植被以及与植被相关的花和传粉者时将出现情况,这时要么在该系统中引入传粉者(例如在杏仁果园加入蜜蜂)要么忍受作物产量的下降。在接下来的部分我们将进一步探索丧失传粉者带来的潜在后果。
5.2 传粉者减少的后果:传粉者多样性,植物多样性,以及内在联系的种子库
由于土地使用方式的改变引起的植物多样性丧失已经被认为是传粉者多样性丧失的主要因素,尽管还有一些其他原因,例如杀虫剂、气候改变、疾病等等(Goulson et al. 2015a,Ollerton et al. 2014,Potts et al. 2010a)。非常重要的是在一个群落中植物物种数量和访花者的数量之间有很强的正相关关系(图4)。通常传粉者要比植物数量多,比如传粉者与植物的平均比率在此设置为2.4(SD=1.5),换句话说,在这个群落中,平均每个植物存在一到两个额外的传粉者,有时候会更多。植物多样性与传粉者多样性的关系也因此有了内在联系:举一个修复项目的例子,我们在群落中增加越多的动物授粉的植物,我们就会找到如预期一样地更多的传粉者。这种关系对保护传粉者有重要的意义,并且是“为了传粉者而种植”全球首创精神和农业管理工作政策的核心内容,但这一定不是需要考虑的唯一因素,因为传粉者也需要繁殖地点和补充资源,包括除了花蜜和花粉以外的食物。
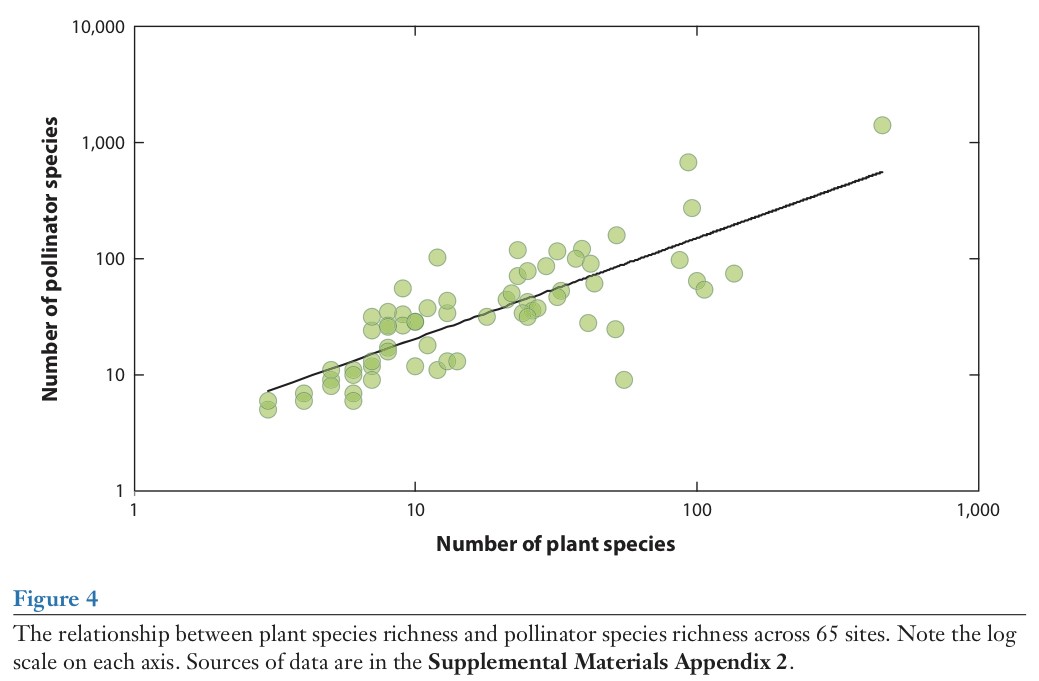
或许是因为植物和传粉者多样性之间的关系太过于明显,所以对它的研究相当少。在生态学中我们经常看到的模型都是尺度依赖型的;例如Hegland & Boeke (2006)利用1.5 ×1.5 m小区域发现植物和传粉者的丰富度整体不具有相关性。与此相反,Ebeling et al. (2008)在一个相当大(20 ×20 m)的草地上同样在德国研究了这种关系。他们发现传粉者物种丰富度与植物多样性有关,然而Steffan-Dewenter & Tscharntke (2001)在同一个国家的农业区域做了连续梯度实验,发现蜜蜂和有花植物物种丰富度之间存在显著正相关关系。图4显著的正相关关系可以被认为是发生在较小空间尺度的多样性的和局部地区的多样性之间关系的全球性表达。
传粉者丰富度和多样性的下降引起了一些研究人员对丧失传粉者隐藏的后果的思考:这会不会导致更大的花粉局限性和种子库的减少,进而影响植物群落的结构?花粉局限性可能由许多因素引起(Knight et al. 2005),其中之一就是某一物种的种群中传粉者多样性较低,就像Gomez et al.(2010)对Erysimum mediohispanicum的研究。与此相关联,Biesmeijeret al. (2006)观察到在英国和荷兰,传粉者和虫媒植物的下降相互关联,这可能表明了原因和结果:较低的传粉者多样性会减少种子的产量进而长期影响植物新种群的产生。然而,用一种实验方法验证这个观点却得到了不同的结论。例如,在Asclepias verticillata (Apocynaceae)斑块中移除熊蜂(Bombus spp.)对这些植物的繁殖并没有明显的影响,因为熊蜂的角色被相同作用的Polistes wasps接替(Hallett et al. 2017),系统内的生态冗余例子见Section 5.1。相反,Brosi & Briggs (2013)发现从局部斑块中移除单一熊蜂物种降低了毛茛科植物飞燕草Delphinium barbeyi (Ranunculaceae)的结实成功率,尽管该斑块中存在可供选择的传粉者。很明显,局部丧失传粉者的后果有特异性,根据涉及的物种和群落环境的不同而发生变化。2012年Albrecht等人用不同的方法发现,在萝卜(Raphanus sativus)群体中增加传粉者将有更大的果实和种子库。在更大的生态学尺度上,2016年Lundgren等人在10个多年生牧草实验中,以群落水平上传粉者的可利用性减少40年,预测了传粉者的减少对籽苗的影响。结果很复杂而且在某种程度上取决于研究的物种,但总的来说传粉者的减少使籽苗丰富度和多样性下降。到目前为止,这是唯一一项研究揭示传粉者衰退和植物物种丰富度或个体丰富度减少之间的直接因果关系。
因此,传粉者减少对农作物的影响更容易预测(如产量),特别是那些具有两性花或其他高度依赖于传粉者的杂交物种。在农作物中有很多传粉者多样性高时产量也增加的例子,例如咖啡(Klein et al. 2003),樱桃(Holzschuh et al. 2012),苹果(Blitzer et al. 2016,Garratt et al. 2014a),以及冬青树和槲寄生(Ollerton et al. 2016)。
但是,传粉者不足影响农作物产量的好例子依旧较少(Aizen et al. 2008),尽管英国苹果产量可能是一个新兴的例子(Garratt et al. 2014b)。但显然传粉者丰富度和多样性的维持对于农业服务和生态功能二者都是至关重要的:等传粉者数量下降到影响农作物产量的水平时再采取保护行为是不可取的。尽管动物授粉的农作物比例较少,但是在富含人类必要营养元素的饮食和经济价值中(IPBES 2016),它们作用重大。一个案例就能说明现代人类社会对动物授粉的依赖程度。咖啡由许多野生昆虫(主要是蜜蜂)授粉(Ngo et al. 2007),而咖啡作为商品就其价值而言仅次于石油,它维持了数百万农民的生计。2016年全球咖啡产量直冲151,624万袋,每袋60kg(Int. Coffee Org. 2017)。一个咖啡豆来自于一个单受精事件,至少一个花粉落在柱头上。平均每个咖啡豆约为0.1g,这就意味着大约每包有600,000个咖啡豆。而2016年的咖啡豆总产量就超过了90万亿个。咖啡的自花传粉概率为50%(Klein et al. 2003),蜜蜂访咖啡花时,可能携带足够的花粉去使每一个咖啡花的卵子受精,所以我们可以把这个数字除以四。尽管如此,全球咖啡产量仍然需要至少22万亿个传粉者去访花。显然,有了数十亿只需要半栖息地和咖啡种植园的蜜蜂,全球咖啡市场才得以生存。
6.总结
自中生代以来,传粉者已经为植物服务了至少1.7亿年,可能比想象的更久。在那个时期,不同传粉者群体的相对重要性也有变化,但整体传粉者的多样性与有花植物平行增加,直到目前,多达350,000个传粉昆虫被描述记载(还有更多的有待于科学发现)。不同分类群体(从属的水平到目的水平)的相对重要性也存在变化,但总体来说,生物多样性很重要,任何一个物种的丢失(无论在什么地理尺度)都应该避免。同时,我们不希望当前的模式一成不变,排除人为干扰过程,地方和全球物种的丢失和增加都将成为自然生物多样性波动的一部分。
目前,对传粉者多样性和重要性的全球模式的理解,以及对不同授粉系统的角色(特别是风与动物授粉相比)的理解具有重要意义,但仍有许多需要探讨的地方。在过去的20年里,科学家对这些问题产生兴趣并开始就这些问题收集全球数据,如在世界上风和动物传粉是如何变化的,作为传粉者不同群体的动物的重要性如何显现,花粉限制模式是怎样的,以及这些模式是如何联系植物性别和交配系统的。
理解传粉者多样性,传粉系统的演化过程,传粉者扮演了什么样的角色,传粉者嵌入生态网络等问题(所有的这些是如何作为生物多样性至关重要的一方面而被保存的)需要更多的观察和实验数据以及监控和详细的调查才能确定一个传粉者多样性下降的画面。现代传粉者的多样性是数百万年来裸子植物和被子植物紧密扩散、协同进化的结果。作为植物多样性的关键延续,传粉者也通过间接地支持大量其他有机体提供巨大的附加价值,这些有机体包括酵母以及花蜜中的其他微生物,花的寄生真菌,盗寄生蜜蜂物种和其他的寄生虫,专一的捕食者和食草动物,食果-食种子动物等等。失去这其中任何一个多样性将是我们地球生物遗产的悲剧性损失。
https://blog.sciencenet.cn/blog-536560-1085804.html
上一篇:高通量识别核基因多信息位点在浅层次系统发育和谱系地理中的应用
下一篇:第二届生物系统学论坛报告人名单