博文
整合古生物学和系统发生学方法研究宏观进化
||
整合古生物学和系统发生学方法研究宏观进化
Integrating Paleontological and Phylogenetic Approaches to Macroevolution
Annual Review of Ecology, Evolution, and Systematics
Vol. 47:189-213 (Volume publication date November 2016)
First published online as a Review in Advance on August 19, 2016
https://doi.org/10.1146/annurev-ecolsys-112414-054207
Gene Hunt1 and Graham Slater2
1Department of Paleobiology, National Museum of Natural History, Smithsonian Institution, Washington, DC 20560; email: hunte@si.edu
2Department of the Geophysical Sciences, University of Chicago, Chicago, Illinois 60637; email: gslater@uchicago.edu
摘要
随着分子系统学的发展和统计模型的发展,系统发生学者现在可以解决传统上一直是古生物学范围内的宏观进化问题。在系统发生和古生物学研究的交叉点上,人们的兴趣集中在三个领域:时间尺度的系统发生、理解性状演变和建立物种多样化模型。长期以来,化石标定对于将系统发生定标为绝对时间至关重要,但是最近的进展同等融合了灭绝类群。模拟和实验研究表明:化石数据可以显著改善关于性状演变的推论,特别是对于具有非均质时间动态的模型以及生物形式并不能代表其较大进化枝幸存者的进化枝中。近年来,关于推断多样化动态的性质和不确定性,也进行了富有成果的跨学科对话。挑战仍然存在,但是现在代表着将这两种关于生命历史的观点整合的兴趣盛行起来。
关键词
1. 引言
传统上,通过研究化石记录来研究大规模的进化动态和趋势(Simpson 1944;Jepsen等,1949;Stanley 1979)。最近生物物种的系统发生数据的爆炸式增长以及系统发生比较方法的同期发展,宏观进化动态由此可以从中推断,这为进化生物学家打开了这一研究领域。在某些情况下,这导致分支动态的解释大相径庭,并且可用于系统发生数据的方法和模型的日益复杂,使人们开始质疑是否需要化石数据(例如,Monroe & Bokma,2010; Venditti等,2011; 贝克等,2015年)。尽管存在这些冲突,古生物学家和生物学家仍在研究同一棵生命树,我们应该通过结合树的帮助,从现在起回顾过去的观点,并通过随着时间的推移的化石记录,来丰富我们对宏观进化的共同理解。尽管已经开始对此观点达成共识(Fritz等,2013;Pennell&Harmon,2013;Benton,2015),但仍然存在实际挑战,目前尚不清楚在哪种情况下,古生物学或系统发生学方法应该最有用。
在这篇综述中,我们试图总结我们认为与系统进化和古生物学方法整合研究宏观进化最相关的领域的现状。我们没有讨论其它地方已广泛评述的方法的机制(例如,O'Meara,2012; Morlon,2014),而是关注系统发生和古生物学共同关注的三个主要领域:将化石信息整合到时间尺度的系统发生中、推断形态进化的速率和方式、了解物种形成和灭绝的动态。最后,我们讨论一些似乎更有望实现更完整整合的领域。
2. 有化石的树
整合系统发生和古生物学研究宏观进化最明显途径是将化石类群插入现存物种的系统发生树中。原则上,这很简单:如果可以为现存和化石类群编码特征,则可以对组合数据集进行分析。但是,挑战比最初看起来要多。在本节中,我们回顾了正在进行的两个主要发展领域:如何将化石物种放入生物物种树中,以及如何对这些树进行时间缩放,以便将其用于下游分析?
2.1. 把化石放到树上
除极少数情况外,形态学是评估化石物种与现生类群关系的唯一手段。传统上,形态学系统学家的默认方法是最大简约法。在简约准则下,我们应该更偏向给定特征矩阵所暗含的特征状态变化次数最少的系统发生假设。Felsenstein(1978)指出:在某些条件下,简约性在统计上是不一致的。这意味着随着数据量的增加,返回错误答案的概率也会增加。当一个或多个分支在同一个变化率下表现出比预期更多的特征状态变化时,最常发生这种假象;并且,由于越来越可能采集到趋同的相似征态而导致这些长分支末端类群相互吸引。化石类群(如果有的话)可以通过分解长分支并告知征态转换的极性,从而在简约分析中起关键作用(Gauthier等,1988;Huelsenbeck,1991)。但是,如果沿基础系统发生内部分支的形态进化速率太快,则无论是否采集化石,简约法在恢复正确拓扑结构方面仍可能做得不好(Wright & Hillis,2014)。
在分子系统学中,长分支通常不能被打破,人们意识到即使在同一个基因内,进化速率在各个位点之间也是异质的(例如,转换比颠换更频繁;第三个密码子位置比第一个或第二个位置进化得更快)。基于更现实的序列进化模型,概率系统发生得以发展。Lewis(2001)基于征态之间变化率相等的马尔可夫转换(M k ,其中k 表示特征状态的数量)模型,提出了形态特征进化的相应模型。概率模型用于形态数据推断系统发生推断已被证明对某些人存在哲学争议(Spencer & Wilberg,2013),但得出的统计结果没有争议。例如,模拟显示,对于由现存和化石类群组成的数据集,当进化速率快或异质时,使用M k 模型进行贝叶斯系统发生推断所产生的拓扑错误百分率要低于简约性(Wright & Hillis,2014)。使用古生物数据集进行基于模型的推理也可以通过根据解剖区域对特征进行分区(Clarke & Middleton,2008)或通过概率分布中的采样率(Wagner,2012 ; Harrison &Larsson,2015)进行改进,就像对分子数据所做的那样。的确,通过提供适当的统计框架来分析在异质过程中进化级联数据集,对形态学数据使用基于模型的推断可以更好地把化石整合到现生类群的分子系统发生中(Bull 等,1993)。最近的工作表明,与化石保存相关的过程可能会非随机地去除特征,从而使特征改变更容易丢失,从而使化石类群看起来比实际的更为原始(请参阅Sansom & Wills(2013)中的 “stemward slippage” )。将简约方法与基于模型的方法用于人工破损的数据进行比较表明,后者在避免茎干滑移方面比简化方法更胜一筹(Pattinson等,2014;Wright & Hillis,2014),尽管需要探索进一步的情况。我们应该更偏向某种方法。
尽管已经做出了很多努力来表明缺失数据本身并不是基于模型的系统发生学中的压倒性问题,但是无论采用哪种推断方法,缺失数据的结构对于整合灭绝和现存生物到同一系统发生中可能都是有问题的。直觉可能表明,现生形态学家将数据收集工作不成比例地集中在解决高阶元的关系上,至少对于哺乳动物而言,这似乎是正确的(Guillerme & Cooper,2015)。这为化石整合到现生类群的系统发生中构成了重大障碍,因为现生类群的完整分子但部分形态数据集提供的信息很少,无法完善仅以形态学表示的化石类群的位置。反过来,这会导致不稳定的拓扑布局(Guillerme & Cooper,2016),并且可能以与整合随机分辨率分支格局相同的方式扭曲下游的宏观进化分析(Rabosky,2015)。拓扑搜索可以是固定的,也可以是严格限制的,以使用没有相关特征数据的推断现存近缘类群来恢复化石类群(Ronquist等,2012;Slater,2013),但这同样可能导致结果有偏见。完整的现生和灭绝类群系统发生的最大障碍可能仍然是需要增加现生生物形态学数据采集以指导化石的放置(Guillerme & Cooper,2015)。
2.2. 化石类群系统发生的时间标定
时间是树推断的信息成分,因此是全证据的一部分,这一观点在古生物学文献中有悠久而争议的历史。我们将不在这里回顾大量文献。在《自然》辩论http://www.nature.com/nature/debates/fossil/中,可以找到一些过时但仍能提供有益信息的观点摘要。取而代之的是,我们认识到可以采用两种主要方法对现生生物和灭绝生物的系统发生进行时间标度:同时分析具有地层数据的特征和事后缩放。
2.2.1. 同时推断。
在系统发生分析中直接组合特征和地层数据的基本论据是,在所有条件都相同的情况下,类群在化石记录中出现的顺序应该告诉我们一些有关系统发生的信息。给定一系列相互竞争的系统发生假说,我们应该选择那些既能最适合性状数据又能适应化石年龄分布共识的假说。这种想法产生了地层债务的概念(Huelsenbeck 1994;Clyde & Fisher 1997),它是一种量化系统发生树隐含的化石记录缺失数量的方法,通常是根据离散的时间段,例如阶段或陆地哺乳动物的年龄(图1)。地层支序学(Fisher,2008)虽然不是严格意义上讲的一种时标方法,但它第一个证实了以下观点:从简约性推论得出的最小长度树,在采用稍长的树并考虑了地层数据之后可能会被拒绝(Clyde & Fisher 1997)。Huelsenbeck & Rannala (1997)提出了一种最大似然方法,仅使用地层信息即可从化石类群的系统发生中推断出物种形成和灭绝时间。Wagner(1998)扩展了该模型:通过基于化石采样强度和灭绝率模型的最大似然方法(Foote 1997a),结合了使用简约评分的参数自举对特征似然逼近。将这种方法应用于Hyaenidae的系统发生中,他获得了系统发生的最大似然估计。与简约分析相比,该系统需要另外11个特征状态变化,但隐含的地层债务减少了39个单位(图1)。
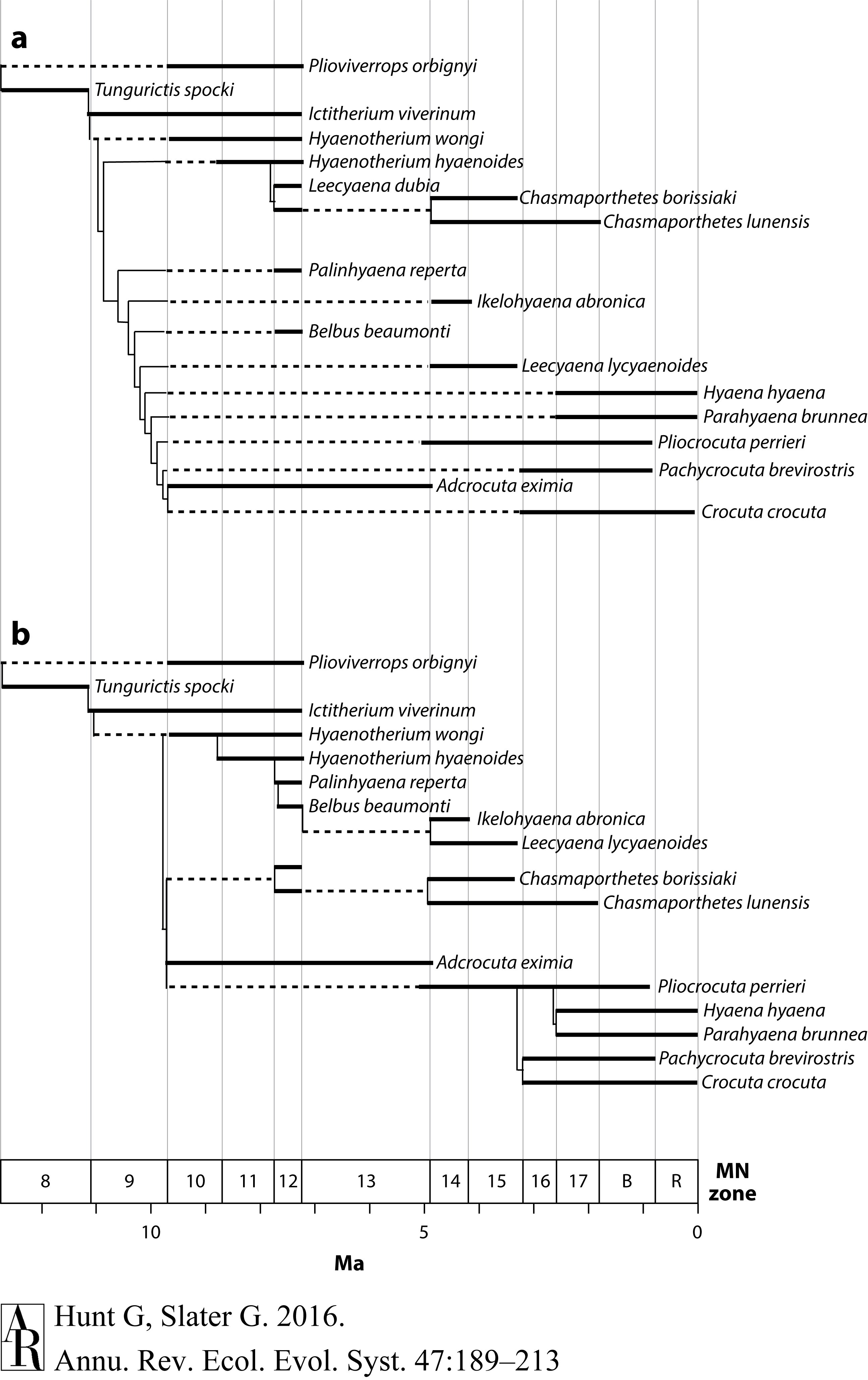
Figure 1 Accounting for stratigraphic debt can result in different optimal tree topologies. The hyaenid phylogeny depicted in panel a is the most parsimonious tree based on analysis of 19 morphological characters for 18 species and results in 47 units of stratigraphic debt after allowing for ancestor–descendant relationships. The tree in panel b is 10 steps longer (50 versus 60 morphologic steps) but has only 11 units of stratigraphic debt, making it the more parsimonious phylogenetic hypothesis overall. Stratigraphic debt is computed as the number of stratigraphic bins, shown here in units of European land mammal (MN) zones, for which a taxon is not observed even though the phylogeny implies its existence. All units are treated as equivalent, so a gap in the relatively long MN13 is considered equivalent to a gap in the much shorter MN14. Abbreviation: Ma, millions of years ago. Figure modified from Wagner (1998) with permission.
直到最近,化石数据在分子系统发生学中的主要作用一直是作为结合现有类群的节点的最低年龄标定。关于结点定年方法的完整说明不在本综述的范围内(请参阅Donoghue & Benton,2007;Ksepka等,2011)。只是要注意,这里的想法是,如果可以确定地将化石分配到特定的进化枝,那么它的年龄为该进化枝的年龄提供了下限。但是,由于进化枝中哪个特定分支与化石相关通常存在很大的不确定性(它可能在进化枝历史的早期或相对较晚就分化了),因此通常使用概率分布来表示化石的合理范围。根据该先验分布和分子数据,使用进化枝年龄较大的方法和数值方法来获取年龄的样本。
最近,新的文献开始出现,就像早期的古生物学方法一样,其重点是通过称为支端定年的过程将化石数据同时整合到系统发生推断和时间标定中(Pyron,2011;Ronquist 等,2012)。支端定年通常假设化石和现生类群的形态特征数据可用,并且它们以与分子数据相同的方式以严格或宽松的时钟方式进化(尽管这不是严格必要的;请参见Heath等(2014)。由于化石类群具有相关的发生时间,因此随后有可能标定形态钟,进而推断出已灭绝和现存进化枝成员的分化时间。尽管支端定年并没有像地层分支法或地层似然法法那样直接惩罚树形拓扑,从而增加了地层债务,但原则上应有相同的效果;错误地将形态相似但地层遥远的类群分组的支端拓扑应该比最小化地层债务的替代拓扑更不可能,因为它们导致分支特定钟速率分布的可能性较小。
支端定年,使用组合的分子和形态学数据集以及单独的形态学数据集(Slater,2015),已应用到各种各样的进化枝上,从哺乳动物(Slater,2013)到植物(Grimm 等,2014;Larson-Johnson,2016)。尽管从理论上讲优于节点标定,但支端定年树的早期迭代往往表现出夸大的分化时间估计值,对于位于树中较深的节点,有时会出现戏剧性和令人难以置信的情况(O'Reilly等,2015)。有些作者通过把具有附加信息的先验年代置于节点来解决此问题(Slater,2013,20),虽然这样的做法很不理想。这种行为的基础仍有待明确判断,但可能取决于先验,用于加权现存类群和化石类群的时标树中分支事件的预期分布,有时在贝叶斯系统发生学中被称为树先验。早期实现支端定年的(例如,Ronquist等,2012年)使用了先验统一树,将分支时间的任何相对分布都视为先验概率相同。众所周知,灭绝往往会导致重建树(即现存类群树)具有更多偏向支端的分化事件(Pybus &Harvey,2000)。在使用化石物种标定现生类群的系统发生时,统一的先验树的权重相等,而我们知道这不太可能(尽管现存物种的化石代表不会以相同的方式影响我们的先验信念)。
Heath 等(2014) 提出了一个替代模型,他们称其为化石生-死(FBD)过程,该过程综合考虑了物种形成、灭绝和采样率。处理化石数据时,必须考虑采样率,因为化石恢复不良会模仿灭绝现象(Foote 1997a,b),从而使生-死模型中未考虑这些参数的参数估计值产生偏差。与使用统一先验树相比,使用FBD过程推断出的系统发生趋向于导致更合理的分化时间估计,尤其是对于有问题的早期分支节点(Grimm等,2014 ; Heath等,2014 ; Arcila等,2015)。模型最近已进一步发展,以允许类群之间从不同的时间段的直接祖先-后代关系的推断(Foote,1996 b ; Gavryushkina等,2014 ,2015)。
支端定年比地层分支和相关方法有一些好处。使用马尔可夫链蒙特卡洛方法可以很容易地在贝叶斯支端定年中适应系统发生不确定性,而地层分支则往往依赖于树形拓扑结构与地层记录的逐步比较(但请参见StrataPhy程序;Marcot & Fox,2008)。尽管非常有前途,但仍然需要进一步考虑。例如,许多古生物学家认为,分支芽式形成(即,新物种的起源于一个祖先,该祖先在物种形成后继续生长)是化石记录中的常见格局(Bapst,2014a),但是使用系统发生方法很难检测到。Gavryushkina等(2015年)表明,如果在分析中将多个在时间上不同的祖先群编码为单独支端,则这种模式可能是可识别的。同样,在整个系统发生中假设化石采样率(或就此而言,物种形成和灭绝动态)一致无疑是幼稚的。FBD模型存在扩展,允许这些关键参数随时间变化,但是更实际的模型也可以考虑采样概率的地理变化(例如,Wagner & Marcot,2013)。这一领域方法开发发展迅速,我们预计在不久的将来会有更多的进步。
2.2.2. 事后缩放
尽管对拓扑和分支长度的组合分析仍然是理想的,但是对于许多数据集而言,这是不切实际的。古生物学家通常缺乏特征矩阵,例如,使用从多个较小的源树组装而成的超树(例如Betancur-R等,2015),或者没有足够的特征数据来生成特征矩阵,但是可以逐个读取大量采样的化石记录并画树(例如,Aze等,2011年)。在这些情况下,事后缩放方法很有用。事后方法以其最简单的形式缩放分支,特定节点的年龄由其最早后裔物种的首次出现时间决定(Norell 1992)。但是,这种简单的方法可能会引起问题。例如,如果属于一个进化枝的最古老化石不处于最早分化的位置(即,它嵌套了若干节点; 图2a ),然导致后一个或多个零长度分支(图2 b)。已经提出了许多不同的解决方案来应对这种情况,例如,可能参考特征变化程度(Ruta等,2006),任意地向所有零长度分支添加少量时间或在零长度分支上重新分配时间(例如Brusatte等,2008年)。。

Figure 2 Sampling can have significant impacts on time-scaling of paleontological trees. (a) Stratigraphic ranges (black bars) for terminal taxa in the underlying phylogeny (gray). Species (sp) 5 has a stratigraphic range that is older than sp 4, even though it is nested one node above it. As a result, (b) scaling the tree based on first appearance dates only (the “basic” method) results in a polytomy for the lower clade. Furthermore, because no stratigraphic data are available for internal edges, the age of the entire clade is reconstructed to be the same as the first appearance of sp 5. These issues can be somewhat overcome by (c) considering speciation, extinction, and sampling rates using the cal3 method. Here, we show four stochastic realizations of branching times. Although some trees (e.g., the bottom row) look similar to those produced via the basic method, others (e.g., the top row) come closer to matching the true set of branching times. These outcomes emphasize the need to sample multiple realizations of branching times using the cal3 approach. Analyses were done using functions in the paleotree package (Bapst 2012); time was simulated in arbitrary units.
Bapst(2013)提出了一种不同的方法,他称其为cal3。该方法尝试根据需要估计物种、灭绝和采样率模型来随机采样分支时间和分支长度。Bapst的方法是逐步进行的。首先根据最古老、最早发生情况,使用上述粗略方法对树进行缩放。然后,该算法从根部开始访问每个节点,并对节点年龄进行采样。然后,对后代分支进行缩放,以扩展到支端,或者扩展到内部节点的随机抽样年龄的最后出现时间。Bapst(2013)使用速率标定模型来估计节点年龄是关键,因为它提供了一个合理的期望,即从一个类群与其姊妹支系的分化到其在化石记录中首次出现之间,我们可能期望缺失了多少进化时间(图2 c)。因此,cal3 方法在本质上与Wagner(1998)相似,因为它不一定要尽量减小分化和首次出现之间的差异。
随着系统发生比较方法(PCMs)越来越多地用于探索仅由灭绝类群组成的大型(> 100类群)树中的宏观进化动态,事后缩放方法可能会变得更加突出。拟合宏观进化模型时,该cal3 方法似乎导致时间缩放的树比简单缩放方法产生更好的结果,但表现并不完美(Bapst,2014A)。在不久的将来,这一领域将得到更多关注。
3. 性状演变
古生物学家和PCM从业者都试图了解性状演变格局、过程和动因。而且,相对于种群遗传学家而言,他们的数据通常具有类似的局限性:时间分辨率粗糙,而与微进化过程直接相关的信息却很少。与所有科学家一样,他们也处理不完整的信息。因各种地质、生物学和采样原因,化石记录出名的不完整,所有这些原因都可能干扰进化推论(Kidwell & Holland,2002)。系统发生专家可获得的信息并未通过地质记录过滤,但信息也不完整。现代多样性通常是不完全采样的,通常是非随机采样(Garamszegi & Moller,2011),当然也缺乏直到今天还没有生存支系的直接证据。
表型性状变型几乎是无穷无尽的,但是在编码上以定性离散状态编码的特征(例如特征的存在/不存在)与那些连续尺度变化的特征(例如体型)之间必须进行重要的实际区分。这些数据类型需要不同的模型和分析程序(O'Meara,2012)。离散状态的分析主要依赖于第2.1节中描述的M k 模型。尽管大多数都是布朗运动(BM)或Ornstein–Uhlenbeck(OU)模型的变体,但连续性状的模型种类繁多。BM是一种无界扩散过程,其漂移参数指定方向变化,变量参数指定扩散速率。通常,PCM专注于非定向进化的特殊情况,其中漂移参数为零,扩散变量控制性状演变的速率。BM的常见细节涉及随时间或跨树分支而改变此速率。OU模型具有类似BM的动态特性,并具有吸引人的最佳效果,可以像橡皮筋一样将性状拉向它。出于评述的目的,我们将各种数据类型的示例混在一起,因为整合古生物学和系统发生方法的好处和挑战是相似的。
3.1. 估计祖先征态
许多早期PCM工作都集中在估计未采样祖先的表型上,以更好地理解进化转变。即使在统一的进化模型,如BM(无方向性漂移)和等速率的M k 模型(Schluter等,1997)下,与这些估计相关的不确定性也可能很广泛。将化石纳入此类分析可以极大地改善祖先状态的估计,因为随着人们试图从采样的类群中追溯到更远的时间,不确定性会增加。灭绝类群在时间上更接近要估计的祖先节点,这些较短的路径长度可以大大减少估计的不确定性(Polly,2001;Finarelli & Flynn,2006)。
当时间异质性或选择性灭绝使进化枝的现生成员无法代表祖先形式时,加入化石的收益将更大。例如,Webster & Purvis(2002)和Finarelli & Flynn(2006)证明了浮游有孔虫和犬形哺乳动物现生成员的祖先体形估计值是不准确的,这是因为每组的体形都有增加的趋势。在后一个示例中,仅对现生犬类动物的分析表明祖先的体型为23千克,而包括化石类群的估计<2千克,与该组中最早的已知化石的体型一致(另见Albert等,2009;Pyron & Burbrink,2012;Slater等,2012a)。最近的另一个例子来自Betancur-R等(2015),他们研究了辐鳍鱼类的祖先栖息地(海洋还是淡水)。仅分析现存生物有利于淡水祖先,但增加了许多化石类群却颠倒了结果,有力地支持了进化枝的原始海洋生境。在这种情况下,现代动物群误导了人们,因为几只具有海洋祖先的基部鱼类在海洋环境中经历了选择性灭绝,现在只限于淡水。
3.2. 拟合进化模型:化石类群的好处
尽管祖先状态估计可以为评估特定的进化场景提供信息(例如,辐鳍鱼起源于哪个栖息地?),但PCM领域已将重点从祖先状态转移到显式性状变化模型上。这种转变强调将进化假设作为指定模型进行评估,然后利用现代统计工具(如似然、信息标准和贝叶斯方法)的能力来拟合和比较观察到的性状分布的竞争性解释。
无论是否结合了化石类群,对性状演变进行系统发生分析的机制几乎没有什么不同。需要一棵树或一组树,通常使用第2节中讨论的一种方法将分支长度按时间单位缩放。终端类群的特征在于其一个或多个性状的值,通常通过参考进化变化合适的模型进行评估。出现的一个复杂情况是,有时实施性状演变方法会假设树是超度量的,将化石整合到现存的类群中时,这种假设会被违反(Slater,2014)。假设一棵超度量树可以简化编程,但是当做出这种假设时,这对于最终用户而言并不总是显而易见的(Slater,2014)。只要可行,就应避免这种做法。
与祖先状态的估计一样,当进化动态随着时间的推移而不一致时,添加化石类群以拟合模型的好处也会不成比例地增加。Slater 等(2012a)在连续性状变化的五个模型下模拟了性状变化:BM、具有趋势的BM、OU、减速演变(也称为早期爆发模型)和加速演变。他们发现,对正确模型的模型支持不受BM和OU模型下添加化石类群的影响,但是通过添加化石类群而大大提高了性能(其预期性状值或进化速率随时间变化)。支持正确模型的倾向增加,不仅仅是因为在分析中增加了更多的类群。当作者将现存类群和化石类群一对一交换时,化石类群的性能优势仍然存在。因此,在每个类群的基础上,化石类群可以比现有类群提供更多有关进化模式的信息(Slater等,2012a)。
连续数据的趋势模型尤其依赖于化石信息以便进行估计。方向性信号是性状值和经历时间之间的系统关系(Hunt,2006)。超尺度树中的端部类群与根节点等距,因此不包含有关趋势强度的信息,至少对于具有趋势的统一BM模型而言(参见3.5节)。当一棵树中存在灭绝类群时,类群年龄的最终范围允许估计趋势,而实际上,即使将几个灭绝类群散布到仅有现存类群的树上,支持趋势模型的趋势也会急剧增加(Slater等,2012a ;有关有或没有化石的离散性状特征的方向性示例,请参阅Klopfstein等,2015)。
试图测试哺乳动物体型增加趋势(Cope法则)的尝试很好地说明了这种效果。古生物学家长期以来一直认为,哺乳动物的支系会随着时间的流逝而变大,这得到了Alroy(1998a)研究来自充分采样的北美化石记录中的1,500多种物种的支持。然而,Monroe & Bokma(2010)在对3,000多种现存哺乳动物物种进行系统发生分析时,几乎没有支持Cope的规则。在该研究中,趋势模型是可以估计的,因为他们的模型允许进化集中在物种形成事件上。这有效地使树成为非超尺度树,因为来自根部到端部,物种形成事件(推断的)数量发生变化。Bokma等(2015)更新了这种方法,纳入了500多种已灭绝哺乳动物物种的体型估计,这些物种被用于设置现存物种树内部节点的先验信息。新的、化石-现代综合分析发现了对Cope规则的压倒性支持。除了化石信息外,这两项研究使用的数据和方法非常相似,这表明几乎所有方向性信号都来自哺乳动物的化石数据。然而,最近两项关于现生类群的研究已经能够得出哺乳动物的祖先体重估计值,即使面对体型趋势也与化石估计值一致(等,2015 ; Puttick & Thomas,2015)。两项研究都允许在树的各个分支上进化速率的变化,这显然允许在没有化石的情况下推断出这种动态(尽管添加化石后,祖先大小估计值附近的置信区间大大缩小;Puttick &Thomas,2015)。
除了表明体型增大的趋势外,Alroy(1998a)的数据汇编还支持传统观点,即白垩纪/古生代生物灭绝后哺乳动物的体型差异显著跃升,大概反映出非鸟类恐龙灭绝后留下的生态位空间的起源脉动。这种效果在视觉上很明显:即使未标记时间轴,也可以在Alroy的图形上找出灭绝边界。尽管如此,对大量现存哺乳动物系统发生的分析,尽管找到了足够的证据表明分支间的速率变化,但没有发现与大灭绝相关的体型进化速率增加的信号(Venditti et al,2011)。与此相反,一个在现存和灭绝的哺乳动物整合个体大小的分析(Slater,2013,20)中发现:强力证据证明进化模式从白垩纪约束(OU)分化转变为到大灭绝后的无界(BM)动态。这很可能是这样一种情况,即远古时代强烈的进化模式的信号被随后的灭绝所侵蚀,因此不易从现存动物区系检测到。
3.3. 物种形成会导致表型变化的脉动吗?
Eldredge & Gould(1972)在现在著名论文中提出,大多数物种在其一生中变化不大,这种现象被称为停滞,而大多数表型变化则与物种形成事件相关。这个间断平衡模型引起了古生物学家(Gingerich 1985)和进化生物学家(Charlesworth等,1982;Pennell等,2014)的争议,这激发了不同领域的科学家收集证据以评估其主张。
尽管收集了数百篇古生物学案例研究来评估间断平衡的停滞部分(Hunt等,2015),但测试物种形成(或分支形成)变化更加困难,因为在化石序列中很少捕获到物种形成。Alan Cheetham及其同事(Cheetham 1986;Jackson&Cheetham 1990)认识到,适当地测试间断平衡需要将祖先-后代序列拼接到系统发生框架中,以便可以在支系内部(遗传的)和支系间(分支发生的)单独估算变化。当作者将这种方法应用于苔藓虫Metrarabdotos 丰富的新近纪化石记录时,他们发现进化变化主要与物种形成有关,并且物种首次出现在化石记录中后便绝大部分经历了停滞。
Bokma(2002)建立了混合的分支形成和遗传形成的进化模型,该模型使间断平衡类似的动态的特征适合于现存物种的系统发生树(另见Bokma,2008;Bokma等,2015)。这种方法需要推断出“遗漏”的物种形成事件,即,在现存树中不可见的分裂,因为灭绝已经修剪了检测它们所需的支系。此外,仅凭现代数据很难区分出遗传形成和分支形成成份。参数估计值的置信区间通常包括与强遗传形成和强分支形成变化均一致的解决方案。化石数据有可能在此起到帮助作用,因为化石种群的祖先-后代序列提供了有关遗传形成变化的信息,该信息不受与分支形成相关的变化所污染。但是,当通过Ostracode进化枝上的系统发生树和化石祖先-后代序列共同估算Bokma模型时(Hunt,2013年),即使在化石约束遗传形成的条件下,性状演变的遗传形成和分支形成成份的相对重要性仍存在相当大的不确定性。
从表型到分子分化,Ezard等(2013)发现,与仅现生系统发生或任何生态或环境因素的相同计数相比,现生和化石有孔大孔虫的系统发生中根部与支端之间的结点数目更好地预测了物种特定的分子进化速率。该结果提供了比仅从现生类群的系统发生获得的假说与分子进化脉动有关的假说更有力且更有说服力的检验(例如,Pagel等,2006)。
3.4. 形态变化差异和早期爆发
在古生物学中出现了差异或形态多样性的研究,以尝试检验经典观点,即随着大类群的辐射,早期支系迅速扩散,从而定居了大多数可用的形态和生态空间,随后的支系则填充了空隙(Simpson,1944; Valentine 1980)。Foote(1990,1991)介绍了随时间量化形态差异的方法,通常计算为多变量变化(或有时作为范围或形态空间占用量,或者作为独特特征组合的数量)。随后进行了许多关于不同进化枝和性状的实证研究,得出了各种各样的差异轨迹(Foote 1997b),其中差异的早期高峰很普遍(Hughes et al,2013)。
差异的早期高峰与性状演变的速率一致,该速率最初较高,但随后下降,但是相同的格局也可能由选择性起源和灭绝,甚至以恒定速率但在有限的形态空间内进化而引起(Foote 1996a)。需要系统发生信息来区分这些相当不同的宏观进化过程。最初,可以通过扩展BM来模拟形态空间的快速探索,以使其速率参数随时间降低(Blomberg et al,2003)。然而,当应用于现存的类群树的汇编时,这种早期爆发模型很少被用来作为解释体型大小或形状进化(Harmon et al,2010)。
是什么解释灭绝和现生之间明显的差异支持了早期爆发模式?模拟表明,检测早期爆发模式的能力可能非常有限,但是当添加化石时会有所改善(Slater & Pennell,2014)。因此,人们可能希望这种模式在现存物种的系统发生中可能不会被发现,特别是如果它们的大小适中。另一个因素可能是系统发生或分类学的研究规模:大多数古生物学差异研究的特征是广泛的高等类群的多样化,但是Harmon等的汇编(2010)收集了相对较小的进化枝。具有更广泛分类学范围的研究偏好早期爆发和类似的模型(例如,Cooper和Purvis,2010),特别是在古生物树中(Wagner,1995;Close等,2015;Hopkins和Smith,2015),尽管爆发是否与分支启动相吻合取决于哪个节点被视为起点。
关于进化速率的早期爆发是罕见的还是普遍的尚无定论,尽管有足够的证据表明进化枝的进化速率存在差异(Eastman等,2011;Rabosky等,2013)。差异在化石比较研究中仍然有用,但最好将其与基于模型并基于系统发生的分析结合起来。对较高级阶元进化枝的研究与重点放在何处而不是是否发生突飞猛进的进化相结合,将是系统发生研究未来探索的一条富有成果的途径。
3.5. 性状的古生物学时间序列
化石信息的基本优势是其时间范围,因此古生物学家定期构建时间序列以了解性状演变的模式也许并不奇怪。当时间序列中的点对应于被解释为具有直接祖先-后代关系的种群时,则时间序列是表示它们之间系统发生依赖性的正确方法。对于这些遗传发生支系,PCM中许多的性状演变相同模型可以采用相似的,基于似然性的方法(Hunt,2006,Hunt等,2015),并将结果直接与系统发生数据进行比较。
与本综述更直接相关的是一些例子,其中时间序列用于总结整个进化枝而不是物种水平支系内的性状演变,特别是趋势(Stanley 1973;McShea 1994,Novack-Gottshall & Lanier,2008;Finarelli & Goswami,2013;Heim等,2015)。尽管时间序列方法可以解释时间间隔之间的时间自相关,但是它们不能正确解释物种之间的系统发生依赖性。而且,时间序列既可以反映支系内的性状微进化,也可以反映相对于焦点性状的物种分化或灭绝造成的物种分选(即物种选择)。例如,体型大小的时间序列可以通过三种机制的任何组合(图3)显示出增加的趋势(Cope规则):(a )构成焦点进化枝的支系中广泛的方向进化,(b )大型物种中优先物种形成,或(c )小型物种中优先灭绝。从时间序列或时间尺度的系统发生中很难区分这些机制,但其它古生物学观察可能会有所帮助。可以通过记录构成进化枝的单个化石物种内的方向性来检测由广泛的遗传形成导致的趋势(例如,Hunt & Roy,2006),并且当起源和物种灭绝不成比例地集中在具有某些性状值的类群中时,就意味着物种选择(Hansen 1982;Finnegan 等,2012)。
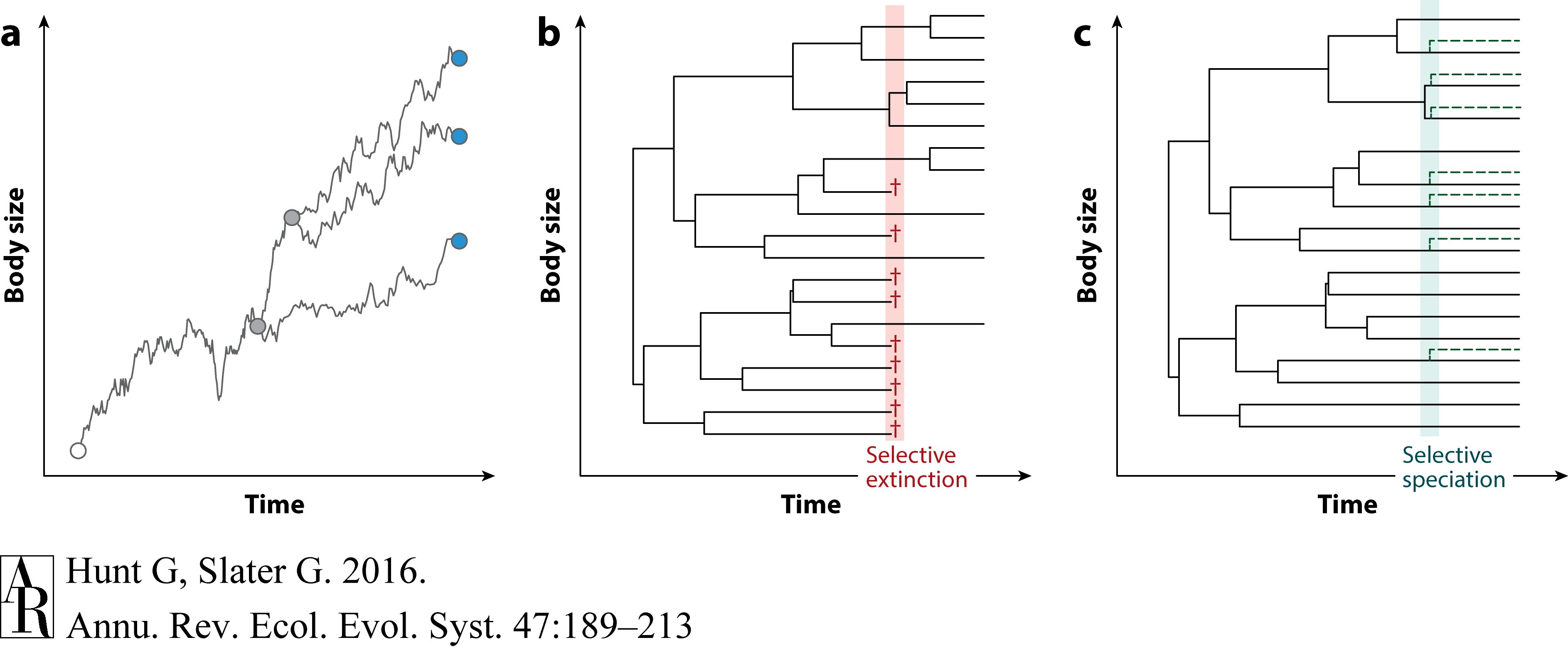
Figure 3 Three different mechanisms that can cause trends are illustrated with examples of Cope's rule, which postulates a pattern of increasing body size over time. In all panels, body size is indicated by the vertical axis and time proceeds from left to right. (a) Trend in a small clade caused by directional evolutionary changes occurring in the clade's constituent species. Lines show the trajectory of trait increase, here modeled as Brownian motion with a trend. Shown are a root node (white), internal nodes (gray), and terminal taxa (blue) at the end of the short simulation. (b) Clade that evolves without extinction until an interval (shaded red) during which a size-selective extinction occurs; daggers mark species going extinct. As extinction has preferentially removed small-bodied species, it increases the mean body size of the clade. (c) Similar scenario to that of panel b, except that size-selective speciation rather than extinction occurs during the same interval (shaded green). The differential proliferation of large-bodied species increases their proportion in the clade, increasing the mean body size of the clade's members. In both panels b and c, changes in body size are shown to occur only at speciation, but the mechanism operates regardless of how changes are apportioned between cladogenesis and anagenesis. Only a single interval of selective speciation or extinction is shown in panels b and c; to generate a sustained trend rather than a one-time shift, such selectivity must be repeated or sustained over time.
4. 多样化
生命已经从数十亿年前的单一支系发展到如今的数百万个物种。这种增加的轨迹,至少对于已经形成的化石形式,一直是古生物学的主要关注点(Simpson 1944;Sepkoski等,1981)。如何在不完整的化石记录的情况下,最好地估算化石多样性(已经在Alroy,2010中进行了综述),我们对此进行了很多研究,在此我们不再重点讨论。相反,我们考虑支持多样性轨迹的起源和灭绝速率,因为从系统发生和古生物学角度来看,这是一个活跃的领域。在性状演变研究中,灭绝和现存的类群基本上以相同的方式使用:作为时间尺度树上的端部类群。在多样化分析中,事实并非如此。可以在系统发生和古生物学方法之间共享这些模型,但是分析的数据和机制相差甚远,值得简要回顾。
4.1. 从化石估算速率
从化石记录中估计起源和灭绝速率的基本观察是一个类群在化石矿床中出现。该类群可能解析到物种水平,但是化石大规模研究中属更常用(Hendricks等,2014;请参见第5.1.1节)。通常,仅在分辨离散时间段(通常与它们相关联的地质阶段或生物地层带)时才知道化石沉积的年龄。这些时段的持续时间通常在数十万到几百万年之间,对化石数据的时间分辨率提出了实际的限制(Bapst,2014b)。较新的高分辨率相关方法有可能产生比以往更高价值的古生物学数据集(例如,Crampton等,2016年)。
多样化的古生物学研究最初是基于观察到类群的第一个和最后一个时间段的计数。开发了方法来估计起源、灭绝和从构成范围端点类群的数据采样率(Foote,2000,2003)。诸如古生物学数据库(http://paleobiodb.org)等基于出现的数据库的日益流行,已将人们的兴趣转移到了在其地层范围内,而不仅仅是范围终点上利用类群发生的方法。这些内部出现很重要,因为它们提供了有关采样强度的直接信息。如果在第一次出现和最后一次出现之间的间隔内未观察到类群,则这种缺失必定是采样失败(除非在化石鉴定和定年时出现错误)。可以通过在许多类群上汇总此信息来估计采样率。这是至关重要的,因为采样异质性会极大地影响观察到的类群发生和从中计算出的比率(例如,Foote,2000;Alroy,2014)。例如,采样间隔较差的时间段可能在最近的间隔中导致类群最后出现的脉冲,从而导致灭绝速率出现虚假峰值。
利用出现数据来限制采样概率的方法包括Alroy(2014)的移动窗口方法,从野生生物学改编的标记捕获(CMR)方法(Liow & Nichols,2010)和最近开发的贝叶斯方法PyRate(Silvestro等,2014年)。所有这些方法都通过联合估计的采样率来估计起源和灭绝,同时考虑化石记录的不完整性。此外,CMR和PyRate提供了丰富的统计框架,可用于跨类群和跨时间的速率变化建模。这些框架允许外部变量影响比率,以便人们可以测试物种形成或灭绝速率是否取决于气候、形态或其它变量。这些功能直到最近才开始在经验研究中得到利用(例如,Liow等,2008;Liow & Finarelli,2014;Pires等,2015;Silvestro等,2015)。
4.2. 从系统发生估计速率
系统发生学从最近共同祖先到如今的物种丰富度,都反映了进化枝的净多样化。不太明显的是,即使树不包含化石类群,时间尺度树的分支时间仍保留有关物种形成和灭绝速率的单独信息(Nee等,1994)。最初,应用于分子系统发生的假设是一个恒定速率的出生死亡过程,该过程均匀地应用于树中的所有分支。该模型被认为是不切实际的,实际上,它通常会产生接近零的灭绝速率(Morlon等,2011年),这与散布着灭绝物种的化石记录不一致。在过去的十年中,为推广这种方法以解释实际的异质性来源而进行的工作激增。由于最近对这些进展进行了综述(Pyron & Burbrink,2013,Morlon,2014),因此在继续探讨这些系统发生方法与化石记录方法之间的相互作用之前,我们仅对它们进行了简要总结。
初步结果表明,在实验系统发生中,多样性下降的信号很普遍。这种格局与物种形成放慢有关,因为辐射越来越多地占据了生态位空间(Phillimore & Price,2008;Rabosky,2013;但另见Moen & Morlon,2014)。捕获了这种减速的度量即γ统计量已被开发出来(Pybus & Harvey,2000),并得到了广泛的应用。随后的方法是明确模拟进化速率的时间变化,如离散向新体制的转移(Rabosky,2006;Stadler,2011)、时间的平滑函数(Rabosky & Lovette,2008b;Morlon et al,2011;Rabosky,2014)、随时间变化的协变量,例如气候代理(Condamine et al,2013)。一些方法还允许进化枝之间的物种形成和灭绝速率发生变化(Alfaro等,2009;Morlon等,2011;Rabosky,2014),随着学者分析越来越大的进化枝,这一点必须越来越重要。最初随时间下降的多样化视为多样性依赖率的指示,而其它一些方法则明确地模拟了多样性依赖(Rabosky &Lovette,2008a;Etienne等,2012)。
另一类模型假设物种形成和灭绝速率受表型性状的影响。原始版本称为二元态物种形成和灭绝(BiSSE),适用于具有两个离散征态的表型性状(Maddison等,2007),每个征态可能与不同的物种形成和/或灭绝速率相关。这种方法很快扩展到适用于多征态和定量性状(FitzJohn,2012)以及性状变化集中在物种形成事件上的情况(Goldberg & Igic,2012;Magnuson-Ford&Otto,2012)。
4.3. 综合多样化分析
即使人们接受通过结合化石和系统发生数据而有很多收获,这样做也是一个挑战,因为很少有类群能结合丰富的化石记录与稳健的分子系统发生,而且许多当前方法都不能分析包含化石类群的系统发生。然而,最近的三项研究比较了古生物学和系统发生方法对同一进化枝的多样化分析。Simpson 等(2011年)研究了过去2亿年的珊瑚多样化,并显示古生物学和系统发生学的曲线具有许多特征,包括侏罗纪中、白垩纪晚期和新近纪的多样化高峰,而在其它方面则速率普遍较低。同样,Cantalapiedra等(2014)对反刍动物的化石记录和系统发生进行了广泛的比较分析。古生物学和系统发生速率的估计值显著相关,尽管它们的对应关系在相对较近的时间间隔内最强,并且仅考虑了现生的冠群成员。这项研究还揭示了古生物学方法(Alroy速率度量与PyRate建模)之间以及从不同研究得出的时间尺度系统发生集之间的重大差异。最后,关于山毛榉目(Fagales:橡树、山毛榉及其近缘类群)的古生物学和系统发生观点较不容易一致(Xing等,2014)。分子系统发生中的支系积累表明,向现在的多样化加速,但化石分析并不支持。这种差异可能是由于进化支活跃的地区化石记录的较低完整性所致。
除了比较系统发生和古生物学方法的个别研究之外,还有一些数据集已从不同角度进行了迭代分析。Quental &Marshall(2010)记录了起源和灭绝的复杂历史,导致化石鲸鱼多样性的蜡化和减弱,他们认为这种格局无法通过当时可用的系统发生方法来恢复。Morlon等(2011年)重新考虑了这个例子,并发现,如果不考虑化石,可以通过考虑系统发生模型来重现鲸鱼多样性下降,该模型的发生率随时间和分支不同而变化(另见Etienne等,2012;Rabosky,2014)。对浮游有孔虫的化石记录进行了充分采样的分析,发现了气候、物种生态学和多样性对多样化影响的证据(Ezard等,2011),随后的系统发生分析证实了这些动态的多样性相关成分(Etienne 等,2012)。
对于化石记录较少的进化枝,即使整合相对较少的化石数据也可以改善对多样化的推论(例如,Pyron &Burbrink,2012)。一种常见的方法是使用系统发生信息来调整观测到的类群的地层范围(Norell 1992)。如果可以假设姊妹类群的年龄相同,则观察到的初次出现的差异可以归因于不完整的化石记录,因此,较早的类群的初次出现可以及时后推,以与其较早的姊妹类群一致。然后,可以将这些调整后的范围用于重建多样性历史记录(对于此方法的某些限制,请参见Lane等,2005;Bapst,2014a)。Jablonski、Roy和其它合作者在一系列论文中将来自双壳类动物丰富的化石记录信息与现代发生数据和部分系统发生数据相结合,以解决各种宏观进化问题,例如气候对纬度多样性梯度的影响(Huang等(2014年)、在系统发生上得到保留并在近缘类群中相似的灭绝率程度(Roy等,2009年)、在系统发生和化石方法中由于灭绝而丧失了进化史(Huang等,2015年)。
5. 整合的挑战和承诺
现在是对宏观进化感兴趣的科学家的好时机。自从1970年代和1980年代间关于间断平衡问题的辩论以来,生物学家和古生物学家之间就一直有持续和富有成效的交流。从事动物区系研究的生物学家正认真地参与化石记录的研究(例如,Pennell & Harmon,2013;Rabosky,2013),系统发生的古生物学家正在迅速采用新开发的基于树的方法。然而,将古生物学和系统发生方法整合到宏观进化过程中仍然面临挑战,并且有希望的脉络尚未被充分挖掘。
5.1. 待解决的挑战
将古生物学和系统发生方法完全整合到宏观进化的进程中存在各种障碍。在本节中,我们重点讨论化石研究与仅现存生物研究之间的数据可比性、方法对数据异质性的强健性和模型违反假设的问题。
5.1.1. 属级和物种级化石数据
化石的多样性分析通常使用属而不是物种作为分析单位。这种选择反映了在属级别工作的实际优势。属比物种更具包容性,因此它们的化石记录更加完整。此外,某些化石群可能难以解析到物种,尤其是在保存方式变化时,并且人们认为属比物种识别更可靠(Hendricks等,2014)。来自模拟(Sepkoski & Kendrick 1993)和实证研究对两种模式都进行了评估(例如,Simpson等,2011;Liow & Finarelli,2014;但请参见Xing等,2014)支持属作为物种有用代理。但是,即使属是物种的良好代表,但在这些分类学水平上,起源和灭绝率的大小将不具可比性。未来的分析发展可能允许在两个层面上转换过程(见Foote,2011),并且欢迎在物种层面开发更多化石数据集,即使此类数据集有其自身的困难(第5.1.3节)。 。
5.1.2. 当违反模型假设时,系统发生比较方法有多脆弱?
一些研究人员已经表示担心,PCM对模型违规可能没有足够的强健性,尤其是在灭绝不可忽略的情况下。Quental &Marshall(2009)证明,只有在某些参数组合下,物种形成和灭绝的情况下,γ统计量才能正确地识别出多样化放缓(另见Liow等,2010;Quental & Marshall,2010)。 Rabosky (2010) 同样表明,多样化率中未建模的异质性可能会影响灭绝的估计(但请参见Beaulieu & O'Meara,2015)。可以通过开发更复杂的模型来处理因违反模型而引发的问题,这些模型可以处理对恒定速率出生-死亡模型的违反(Morlon,2014)。但是,现实可以通过多种方式偏离简单的多样化模型,要评估所有这些,尤其是结合起来的情况,这是一个挑战。最近,对于具有离散类别变量的方法(包括上面讨论的BiSSE系列中的方法),有关PCM强健性的其它问题也浮出水面。这些方法在某些情况下可能会表现很差,至少部分是由于模型违规(Maddison &FitzJohn,2015;Rabosky & Goldberg,2015)。此外,一些作者对连续性状模型对拓扑、分支长度和性状测量中的错误的敏感性表示关注,所有这些都会导致性状中的系统发生信号减少以及对OU模型的强大但不正确的支持(Cooper等,2016)。在基于古生物学系统发生的研究中,这尤其令人关注,粗放解析的树或时间尺度不适当的树正好显示了这种模式(Bapst,2014a;Soul & Friedman,2015)。这里可能需要反复进行方法开发和通过模拟进行测试。
5.1.3. 古生物学方法有多强健?
许多类似的关注点也适用于古生物学方法,尽管它们对于某些会引起现生分析问题的违规行为可能是有力的(Liow et al,2010)。随着时间的变异,在古生物学研究中几乎总是明确地解决,但是化石恢复的时间变异仍然是一个值得关注的问题(例如,Foote,2000;Alroy,2014)。可以使用现有方法处理不同类群之间的差异,但不能始终对其进行评估。原则上,适用于出现数据的现有方法(Alroy指标、CMR和PyRate)可以解释这些异质性。但是它们才刚刚开始被广泛使用,并且将受益于对不同类型的处理变化的性能模拟测试。对于古生物树事后缩放的方法可能引起人们的关注,该方法仅在最近才探索了在已知条件下的性能(Bapst,2014a)。
除了模型假设的问题外,大规模的古生物学数据集引起了人们对数据质量和一致性的重要关注。此类分析通常依赖于古生物学数据库或类似的公共资源。尽管无疑是该学科的福音,但此类存储库托管的数据可能非常不同。除了通常对时间和空间覆盖范围的关注之外,鉴定质量可能非常复杂。理想情况下,分类学专家会审核和统一特定研究的鉴定,尽管这在广泛的时间、空间和分类学尺度上都是艰巨的任务。为了评估特定问题的严重性(Plotnick & Wagner,2006),还应该对被误鉴定污染的数据进行模拟。
5.2. 整合的进一步承诺
建立允许化石和现代数据无缝地为宏观进化推断做出贡献的框架方面已取得了很大进展。如本节中所述,通过在两个学科之间更好地反馈有关数据和模型的信息,以及通过可以处理不理想数据集的方法的额外开发,可能会获得更大的收益。
5.2.1. 交叉传粉系统发生和古生物学模型
尽管多样化的模型在系统发生和古生物学方法中是共享的,但这两个领域通常强调不同种类的变异。这可能很重要,因为一门学科的经验发现会影响另一门学科的建模选择,反之亦然。例如,古生物学家有时会发现证据表明起源和灭绝率在各个群体和时间间隔之间是相关的(例如,Stanley 1990;Xing等,2014)。存在相关性不是完美的例外-否则就不会有差异多样化-但这种相关性可能是系统发生建模方法中当前未集成的一般特征。
另一个示例涉及如何对时间变异建模。古生物学家经常关注速率的间隔变化,这反映了该领域对大规模灭绝和地球历史上其它关键间隔发生的事情的兴趣。相比之下,PCM更关注速率变化的平滑曲线。每种方法都有其优点,但是将每个观点导出到相对的领域可能会有好处。古生物学研究经常揭示出速率的时间序列,这些速率看起来像是波动,偶尔会发生大量的起源和灭绝(例如,Alroy 1998b;Foote,2003;Peters & Ausich,2008),这种动态在大规模灭绝的有限建模之外的系统发生文献中并未得到太多关注(Stadler,2011;Condamine等,2013)。另一方面,古生物学对间隔间变化的关注导致了高度参数化的模型,通常每个间隔都有单独的起源、灭绝和保存速率,尽管对于PyRate方法而言,情况并非如此。对于某些数据集和问题,使用带有较少参数的简单模型可能会改善参数估计并澄清解释。
5.2.2. 环境变量输入模型
通常认为宏观进化过程可能取决于气候等环境因素。最近,Condamine等(2013年)开发了一个模型,其中物种形成和灭绝速率随环境条件变化而变化,而环境条件本身随时间变化。他们使用这种方法来拟合一个模型,其中鲸鱼的物种形成率受温度的影响,而温度是全球气候的替代指标。这种方法(也可以使用CMR和PyRate对仅化石数据实施),从而可以进行以前不可能的综合分析。对环境驱动力的关注还促使系统发生学者和地球科学家在选择和解释地质代理方面进行合作。
5.2.3. 化石记录差和系统发生不完整的方法
Bapst(2014b)强调说,不同类群的化石记录在促成宏观进化推断方面可能存在质的差异。此综述中,古生物学与PCM完全整合的示例往往来自具有良好系统发生数据和丰富化石记录的类群,例如哺乳动物。当今大多数现生进化枝都有稀疏的记录,因此可以利用更适量的化石数据的方法有很大的潜力。对于性状演变,Slater等(2012a)开发了一种方法,该方法将化石表型作为先验整合在现生树的内部节点上。这种方法与常见的情况相吻合,在这种情况下,已知一个或几个化石,但由于不完整而无法正式纳入系统发生分析中。另外,从不完全解析的系统发生的进化枝丰富度推断多样化动态的方法可以很容易地扩展到包含古生物(Alfaro等,2009;Etienne & Apol,2009;Slater等,2012b)。尽管总多样性可以通过不完整的化石记录来估算(例如,Liow & Nichols,2010),但是这样的估算可能是非常不确定的。因此,人们可能希望将化石多样性作为最低估计,或者取而代之的是利用不同时间段或生物分类中的相对多样性来约束多样化历史。
许多化石群有丰富的记录,但很少受到系统发生的关注。在这种情况下,以祖先-后代种对形式存在的家系信息(Alroy 1998a)可以帮助性状演变分析,使用未完全解析的树(上文已指出)或分类学与地层学相结合作为系统发生信息的来源方法也可以帮助分析性状(Soul & Friedman,2015)。
总结要点
系统发生比较方法所采用的模型日益成熟,使得现存物种的系统发生成为了解宏观进化过程,甚至是遥远过去发生过程的可行来源。随着这些发展,越来越多的动力将系统发生见解与化石记录中的见解进行比较和整合。
1. 整合化石和系统发生数据的最基本方法是将现存和灭绝的类群纳入合并的时间尺度树中。最近开发的方法允许将灭绝类群作为定年端部类群(即,通过支端定年)与现存类群同等处理。对于由灭绝类群组成的树,各种事后树缩放可用于将枝状图转换为时间尺度的系统发生树。
2. 通过添加适量的化石数据可以改善祖先性状估计、参数估计和模型推断,尤其是对于随时间变化的进化动态。
3. 尽管较早的推断多样化动态的系统发生方法通常无法得出合理的参数估计值,尤其是对于灭绝而言,但面对恒定速率的生死模型,最近的方法却表现得更好。这些分析在某些情况下重现了化石记录独立的已知格局,这表明它们可有效地应用于化石记录较差的类群。
4. 整合宏观进化的重大挑战仍然存在,包括评估系统发生和古生物学方法的稳健性,使每个学科的经验结果反馈到另一个学科的建模选择,以及进一步开发利用环境变化的地质记录的方法。
https://blog.sciencenet.cn/blog-536560-1216537.html
上一篇:地球生命如何开始的秘密
下一篇:人类世动物区系丧失