博文
Paired Ct. Transformed into Tf. by crude enzyme from tleaves
||
Investigation of Paired Catechins Transformed into Theaflavins by Crude Enzyme of Fresh Tea Leaves (Camellia sinensis)
Tu Yun-fei
Hangzhou institute of tea, China COOP, Hangzhou 310016, China.
Graphical Abstract
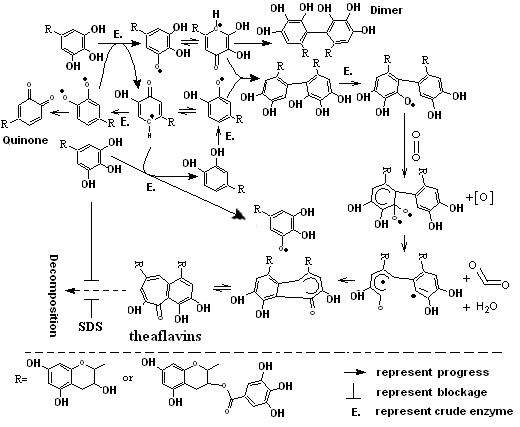
Proposed scheme pathway of theaflavins biosynthesis and EGCG, EGC or SDS play positively role on the theaflavins stability.
ABSTRACT
Background: For elucidating the content of theaflavins in Black tea less than theoretical value, the detailed process of theaflavins transformation with enzymatic reaction was investigated with catalytic model in vitro. Results: the speed of consumption of each catechin catalyzed independently by enzyme was followed as EGCG~ECG>EGC>EC. EGC or EGCG in the medium can keep the formed theaflavins from decomposition and decrease the speed of EC or ECG consumption by sacrifice themselves. Existed epicatechins cut down Theaflavins while epigallocatechins lower to undetectable level. Moreover, same classified catechins can compete with each other for reacting with corresponding catechin. The order of enzymatic reaction of each catechin in pairs can also influence the yield of theaflavins. And the formed micelle system of SDS can help the theaflavins decomposition especially when its concentration over the critical point. Conclusion: priority with oxidation of EGC or EGCG in paired Catechins was in favor of theaflavins biosynthesis, but formed theaflavins were further oxidized along with EC or ECG enzymatic catalysis while EGC or EGCG disappeared in the medium. Surfactant of Sodium Dodecylsulfate could block the theaflavins degradation via the segregation effect of the lyophobic core of micelle with aqueous enzyme.
1. Introduction
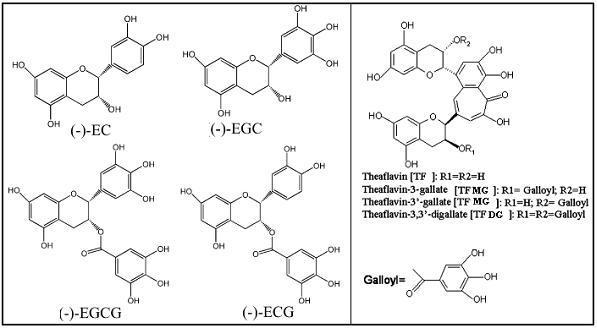
Catechins in pairs including (-)-epicatechin (EC) with (-)-epigallocatechin (EGC), (-)-epicatechin (EC) with (-)-epigallocatechin-3-gallate (EGCG), (-)-epicatechin-3-gallate (ECG) with (-)-epigallocatechin (EGC), or (-)-epicatechin-3-gallate (ECG) with (-)-epigallocatechin-3-gallate (EGCG) can be converted into theaflavins (namely theaflavin (TF), theaflavin-3-gallate (TF-3), theaflavin-3¢-gallate (TF-3¢), or theaflavin-3,3¢-gallate (TF-D-G)) by polyphenol oxidase under aerobic condition or peroxide oxidase conjunction with hydrogen peroxide during the “roll” and “fermentation” process of withered fresh tea leaves(Camelliasinensis (L.), O. Kuntze) manufactured into Black tea[1,2]. Theaflavins as the orange reddish pigments in Black tea were first confirmed by Roberts[3] in 1957 and there have been accumulative evidences demonstrated their positive effects on health such as anti-oxidants[4], anti-cancer[5-11] and cholesterol-lowering effect[12]. Thus, high content of theaflavins in Black tea stand for good quality, flavor and pharmaceutical benefits to humans. And interestingly, there would be 6 %~8 % (w/w) theaflavins formed in Black tea based on theoretical statistics of about equivalent molar ratio between EGC and EGCG and between EC and ECG and of 3~4 fold molecule of EGC plus EGCG more than EC plus ECG in fresh tea leaves[2,13], however, the practical content of theaflavins was less than 2 % (w/w)[2].
The preferable content of theaflavins in Black tea wasn’t still solved well recently, even though the reaction parameters[14-19] such as temperature, pH value and time were investigated and optimized in the model fermentation of aqueous solution in vitro. One of the reasons was ascribed to the fact that the processes of enzymatic oxidation of EC accompany theaflavins consumption by partially purified polyphenol oxidase, which was hardly being controlled in the aqueous solution simultaneously[20, 21]. And EC can promote the oxidative rate of EGC as they coexisted in aqueous solution; while EGC decreased the oxidative rate of EC and play stabilization role on theaflavin content[21] by sacrificing itself. Therefore, EGCwas possessed of positive correlation with the quality of Black tea[22], and surplus of EC or ECG existed in Black tea[2, 23] may result in the current situation. In this paper, we explore the enzymatic mechanism from a new perspective, and the chemicals of hydrogen peroxide, reducing agents and Sodium dodecylsulfate were selected to investigate their effects in the enzymatic process.
2. Experimental
Materials
Standards of catechins (EGCG,ECG,EGC,EC,purity ³ 98 %) were purchased from Shanghai Yousi reagent Co., Ltd. (Shanghai, China). Theaflavins were prepared by the description method[24] in our laboratory,Polyvinylpolypyrrolidone (PVPP,Pharmaceutical grade) was purchased from Boai New Kaiyuan Pharmacy Co.,Ltd. (Henan, China). Nicotinamide adenine dinucleotide phosphate (NADPH), Nicotinamide adenine dinucleotide (NADH), and Dithiothreitol (DTT) (10 mg/ml, purity ≥ 99 %) were purchased from beyotime institute of biotechnology (Jiangsu, China). Vitamin C (Vc, purity ≥ 99.7 %) was obtained from Guangdong Guanghua Chemical Factory Co., Ltd. (Gangzhou, China). Sephadex LH-20 (25-100 mm) was bought from GE Healthcare Bio-Science AB (Sweden). Pure water was got from Hangzhou Wahaha Group Co., Ltd. (Hangzhou, China). Other reagents and chemicals used were of analytical grade and obtained locally. Fresh tea leaves (Jin-Xuan Cultivar, one shoot with two or three leaves) were harvested on April-2011 year, and stored at refrigerator (-30℃) before use.
Crude enzyme preparation and enzymatic condition
3 g fresh tea leaves plus 5 g PVPP was homogenized with 80ml pre-cooled PBS (pH5.6, 100 mM) for 5 min and then filtered through 8 layers of gauze for further centrifugation (4 ℃ for 50 min) at 5500 rpm (Sigma3K15,rotator 11133), and the supernatant as the crude enzyme was transferred into a new tube (stored at -30℃ before use). Aliquot of 0.4 ml crude enzyme in conjunction with a few of standard stock solutions of individual catechins (prepared in PBS, pH5.6, 100 mM) in 10 ml capacity of the circular bottom of graduated glass tube with 0.1 ml graduation was marked to 1.5 ml by PBS (pH5.6, 100 mM) and vigorously stirred on a Vortex mixer (Shanghai Huxi Analysis Instrument Factory Co., Ltd.) before subsequent reaction within water bath at 37 ℃. When the enzymatic time attained, 10 mL of the medium was transferred instantly (<1 min) into HPLC system for quantification of catechins and theaflavins.
Quantification of catechins and theaflavins
The HPLC system consisted of a Waters 1525 binary pump, a 717 plus auto sample injector, a Waters 2487 dual wavelength detector and a reversed phase C18 column (Waters SymmetryTM, Hypersil-C18, 150 mm´i.d.4.6 mm, 5mm) within a column oven (35 ℃), and all the system was controlled by Breeze software. The gradient elution at a flow rate of 1 ml/min followed by description method[25] with some modification: phase A (pure water including 5 % ethyl acetate and 2 % acetic acid) maintained for 2 min, then over 2 min the mobile phase A containing linear increment of phase B (pure acetonitrile) from 0 % to 25 % (v/v) and back to phase A after hold 10 min of this composition. The elution system should be equilibrated for 10 min by phase A before next injection (10 mL). Detection was performed at the optimum violent wavelength (lmax=278 nm). And the acquired curves of standard catechins and theaflavins (5~40 mg/L) were: Y[EGC] = 2896.7 ´ C[EGC] + 2641.3, R2 = 0.9985; Y[EC] = 7183.9 ´ C[EC] - 2317.1, R2 = 0.9995; Y[EGCG] = 18333 ´ C[EGCG] - 5935.3, R2 = 0.9996; Y[ECG] = 22591 ´ C[ECG] - 8854, R2 = 0.9999; Y[TF] = 13621 ´ C[TF] - 5332.5, R2 = 0.9994; Y[TF-3’] = 7488.1 ´ C[TF-3’]- 1265.6, R2 = 0.9999; Y[TF-3] = 4552.4 ´ C[TF-3] - 2469,R2 = 0.999; Y[TF-D-G] = 14502 ´ C[TF-D-G] - 19274, R2 = 0.9964, where Y represent µv×Sec, C represent mg/L, respectively.
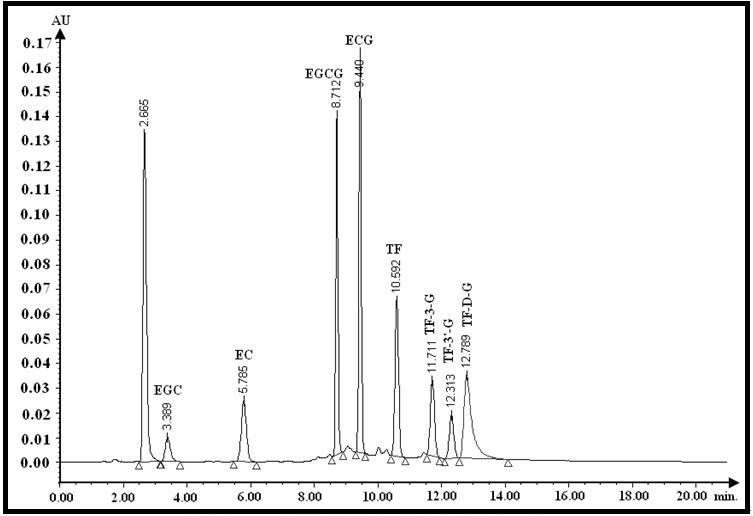
Data processing
Data obtained were expressed as mean ± SD (n≥2). A one-way student’s t test for evaluating the average value as compared with control was performed via office Excel2003 software. A value of p<0.05 was considered to be different significantly.
3. Results
Variation of Catechins and theaflavins in the enzymatic progress
As depicted in Fig. 1.-A, the mole concentration of each catechin was decreased along with time progress and the change rate of each catechin was EGCG~ECG>EGC>EC which wasn’t influenced by catalase (data not shown) when each catechin was catalyzed by crude enzyme independently; And the color of enzymatic solution observed referred with EGC or EGCG was unchangeable all the times and with EC or ECG changed from slightly yellow to deep yellow. Much of evidences have confirmed that a pair of Catechins on B-ring containing ortho-dihydroxyphenyl- and vic-trihydroxyphenyl- group simultaneously can be transferred into corresponding theaflavins by tea polyphenol oxidase or peroxide. Thus, theaflavins except for TF-D-G (only 62 %) were generated gradually as the progressive reaction which were approximately equivalent to the consumed epicatechins in mole such as EC or ECG in conjunction with epigallocatechins in Fig. 1.-B~E before 9 min. The slower increment of TF-D-G than others may be attributed to the relatively heavy galloyl group of EGCG and ECG. Interestingly, we found that the medium was changed from yellow to purple[26] while the enzymatic reaction attained to 7 min which was confirmed with previous experiment using banana homogenate[21] with this paired catechins of EGC and EC. Furthermore, the appeared time of purple prolonged for 10 min when we changed the volume of crude enzyme from 0.4ml to 0.2 ml in the medium. And other groups of paired catechins were only possessed of changeable yellow.
The data additionally revealed (1) EGC or EGCG existed in the aqueous solution not only protect the generated theaflavins un-affected, but also decrease the consumed rate of EC or ECG; (2) EC or ECG accelerate the consumed rate of EGCG or EGC and cut down Theaflavins while epigallocatechins such as EGC[20] or EGCG disappeared in the medium; and (3) The degenerated percentage of theaflavins was TF (76.56 %, t-test, p < 0.05) > TF-3(69.54 %, t-test, p < 0.05) > TF-3(66.71 %, t-test, p < 0.05) > TF-D-G (37.28 %, t-test, p < 0.05) as comparing with 9 min and 25 min. And over amount of EGC or EGCG co-existence in the enzymatic system resist the decomposition of theaflavins effectively in Fig. 1.-F and G, but the decreased tendency can hardly be blocked completely during the enzymatic process. Moreover, content of generated theaflavin such as TF-D-G was positively correlated with any catechin increment in the paired system (Fig. 2.-H). We also observed that same classified catechins can compete with each other for reacting with corresponding catechin, such as group of TF3’ (11.17 %) with TF (-29.63 %), TF3 (17.93 %) with TF (-21.51 %), and TF3’ (83.15 %) with TF-D-G (46.12 %) should be ascribed to increment of the two times of the content of corresponding catechin in Fig. 2.-I. In short time observation, pure theaflavins can’t be influenced by the crude enzyme in the system, and addition of EGCG or EGC also didn’t participate in the oxidation reaction (data not shown).
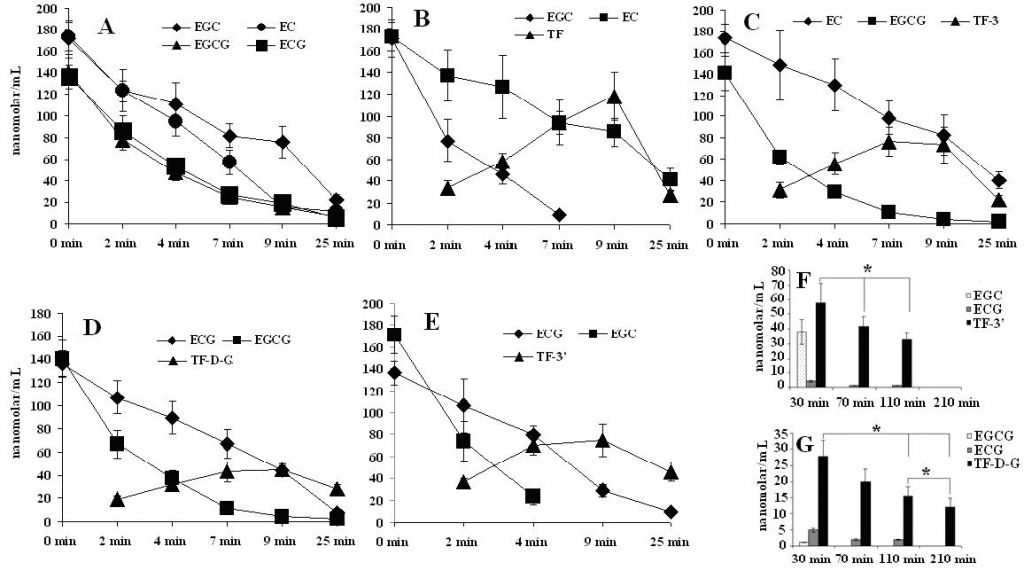
Fig. 1. Molar variation of each catechin and theaflavin with enzymatic process
Note: A represent change of catechins (nanomolar/L) catalyzed by crude enzyme independently; B~E: represent change of TF with paired catechins of EGC and EC, TF-3 with paired catechins of EC and EGCG, TF-D-G with paired catechins of ECG and EGCG, and TF-3’ with paired catechins of ECG and EGC, respectively; F and G represent change of TF-3’ with paired catechins of EGC and ECG and TF-D-G with paired catechins of EGCG and ECG, respectively, based on the initial concentration of 10 folds of EGC or EGCG to ECG (79.12 nanomolar/mL) in the medium. A-EGCG/EGC represent white with slightly purplish precipitation appeared with EGCG or EGC independently when placed the enzymatic medium overnight at room temperature;B-1 represent primary color of EGC and EC enzymatic reaction solution; B-2 represent the purple color appeared when EGC and EC enzymatic reaction attained to 7 min; F-1 and G-1 represent slightly orange precipitation of F and G in when placed the enzymatic medium overnight at room temperature, respectively. * represent p < 0.05.
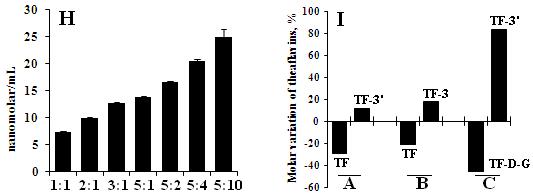
Fig. 2.Molar variation of theaflavins during alterable catechins
Note: H represent molar variation of theaflavins during alterable catechins of EGCG to ECG from 1:1 to 5:10, and 1:1 means 30.72 ± 0.11 of EGCG to 46.32 ± 0.73 nanomolar/mL; A, B, C in Fig. 2.-I represent the mole ratio of EGC: EC: ECG, EGC: EC: EGCG and EGCG: ECG: EGC changed from 1:1:0.5 to 1:1:1, respectively. And 7 min of enzymatic time was carried out in whole experiment.
Order of enzymatic reaction of catechins influence on theaflavins biosynthesis
For elucidating whether oxidized catechins participated in the theaflavins generation, the order with respect to enzymatic reaction of catechins was investigated in the enzymatic process. The results (Fig. 3.) clearly illustrated that more theaflavins except for TF-D-G would be generated in the enzymatic system if epigallocatechin such as EGCG or EGC catalyzed prior to epicatechins than the reversed catalyzed order of the group of paired catechins. And this order result in three times the content of TF and TF3’ as comparing within Fig. 3.-(3) to 3-(1).
On the other hand, if epigallocatechins or epicatechins catalyzed independently and subsequently combined with each other, there was only half amount of detectable TF3¢ in the medium when comparing within Fig. 3.-(3) and deep black precipitation appeared in bottom referred with paired catechins of ECG or EGC via overnight placement at room temperature. Other orders such as the crude enzyme was deactivated after epicatechins or epigallocatechins oxidized firstly or simultaneously in the process could hardly generate the theaflavin. Furthermore, as comparing within Fig. 3.- (8), the theaflavins yield in each group such as Fig. 3.-(1) ~ (7) was less than in Fig. 3.- (8). Thus, over catalyzed catechins especially of epicatechins prior to the paired catechins by crude enzyme may induce down-regulation of theaflavins biosynthesis such as the result of Fig. 3.-(10).

Fig. 3. Effect of order of catalyzed catechins on the yield of theaflavins
Note: (1) ~ (7) represent EC or ECG catalyzed for 9 min and then plus EGC or EGCG for 7 min; EC or ECG catalyzed for 9 min followed by deactivate enzyme with boiling water, and then plus EGC or EGCG for 7 min; EGC or EGCG catalyzed for 9 min and then plus EC or ECG for 7 min; EGC catalyzed for 9 min followed by deactivate enzyme with boiling water, and then plus EC for 7 min; ECG and EGC or EGCG catalyzed 9 min, respectively, and combined instantly for 7 min; ECG and EGC or EGCG catalyzed 9 min, respectively, followed by deactivate enzyme with boiling water, and combined instantly for 7 min; ECG and EGC or EGCG catalyzed 9 min, respectively, followed by deactivate enzyme with boiling water, and combined instantly plus 0.4 ml crude enzyme for 7 min, respectively, respectively. N.D. represent not detectable. And the initial concentration of EC, EGC, ECG and EGCG in medium was 155.31 ± 13.94, 166.84 ± 18.84, 138.62 ± 21.09, 125.36 ± 17.70 nanomolar/L, respectively. (8) Concentration of generated theaflavins in the medium while epicatechins and epigallocatechins existed in the aqueous solution simultaneously. (9) ~ (12) represent EGCG plus 0 min, 1 min, 3 min, 5 min, respectively; (13) ~ (15) represent ECG plus 1 min, 3 min, 5 min, respectively, and the initial concentration of EGCG and ECG in medium was 61.43 ± 0.07 (EGCG), 24.19 ± 1.19 (ECG) nanomolar/L, respectively.
Effects of H2O2, reducing agents and SDS on the enzymatic process
Previous report[27] indicated that enhanced level of theaflavins was attributed by lowering the pH value, which may result in the decreased level of hydrogen peroxide[28] in the medium. And here we carried out the experiment in-vitro by adding hydrogen peroxide in the range of 0~714 mM in the enzymatic medium. The results (Fig. 4.) illustrated that hydrogen peroxide participated in the enzymatic reaction was complex. It can keep the catechins decomposition especially with the maximum concentration more than one time as comparing with its absence from enzymatic reaction. And interestingly, the concentration of theaflavin taken on increased tendency firstly as the concentration of hydrogen peroxide was low and then sharply decreased with its increasing proportion.
In addition to hydrogen peroxide, Nicotinamide adenine dinucleotide phosphate (NADPH), Nicotinamide adenine dinucleotide (NADH), Vitamine C (Vc) and Dithiothreitol (DTT) as the reducing agents were selected to investigate their effects in the enzymatic reaction. The data showed that reducing agents added at the point of 0 min, 1 min, 3 min and 5 min can keep the content of catechins especially EGCG with the two latter agents during the 7 min course of enzymatic reaction. However, the formation of theaflavin was limited gradually as short time as the reducing agents addition. With the effect of co-existence of Vc or DTT added at the point of 0 min, the percentage of theaflavin was only 18 % and 27% respectively as comparing with control; And after from 7 min to 60 min, we observed the concentration of catechins and theaflavin were changed very little in the case of DTT which may be as an effective inhibitor of enzyme. In contrast, EGCG, ECG with TF-D-G were nearly disappeared in the medium after 1hour with the addition of Vc which can be consumed by the enzyme prior to catechins[14].
Furthermore, the concentration of catechins such as EGCG and ECG, which were decreased gradually with the prolonged time as comparing with the total reaction time of 7 min, 150 min and 300 min, were positively proportional to the concentration of SDS (Sodium dodecylsulfate) in the aqueous solution. The concentration of TF-D-G was elevated by the increment of SDS before 1 mmol/mL and then decreased when the concentration over the critical micelle concentration[29] (1.25 mmol/mL) in the case of 7 min, 150 min and 300 min; but the turning point of TF-D-G in the case of 450 min was 8 mmol/mL. As SDS of 1 mmol/ml near the critical concentration, the consumed ECG can convert into TF-D-G over 95% in 7 min which maintained more than 2 h; then decreased by 10 % from 150 min to 300 min and 35 % from 300 min to 450 min with advanced reaction time. In absence of SDS, the theaflavin of TF-D-G can be formed quickly, which concentration was no detectable concentration was formed within 7 min in reversed aspect.
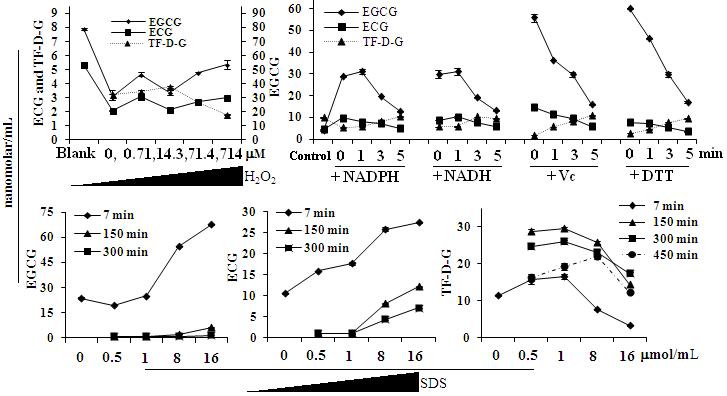
Fig. 4.Variation of Catechins and theaflavins by hydrogen peroxide, reducing agents and SDS
Note: the concentration of NADPH, NADH, Vc and DTT was controlled as 0.5mg/ml. “Blank” means the initial concentration of EGCG and ECG in the absence of enzyme, “Control” means the initial concentration of EGCG and ECG was 61.43 ± 0.07 and 24.19 ± 1.19 nanomolar/ml, respectively. And the initial concentration of EGCG and ECG was 92.65 ± 0.73 and 30.72 ± 0.11 nanomolar/ml, respectively, in the SDS system.
4. Discussion
Theaflavins as the quality index of Black tea have received more and more attention from all over the world because of their benefit effects[4-12] and special flavor[2, 3]. However, the cause of low content of theaflavins in Black tea leaves wasn’t elucidated well. Bajaj [20] found that theaflavins can be decomposed by the progress of EC oxidation in the model fermentation and prevented in the presence of EGC. Furthermore, EC had the ability to accelerate the oxidation rate of EGC in the aerobic fermentation, which was met well with our obtained experimental results. With respect to each catechin oxidized by enzyme catalysis independently, there was no detectable corresponding theaflavin beside from the formation of catechin quinone or di/trimeric catechin such as theasinensis[30] in the aqueous. Furthermore, simple catechins such as EC or EGC in banana homogenate can form 2¢-Glutathionyl-(-)-epicatechin, 2¢-Glutathionyl-(-)-epigallocatechin or 5¢-Glutathionyl-(-)-epicatechin in the presence of glutathione21. And those progressive results combined with our current experiments of order arrangement of Catechins catalyzation and reducing agents effects suggested that the existence of the formed catechins free-radical[31-33] (Fig. 5.) may involved in the process of theaflavins biosynthesis. Finally, in addition to the respect of rate of theaflavin formation in our experiment, the results didn’t support the proposed mechanism of theaflavins biosynthesis based on catechin quinone[21].
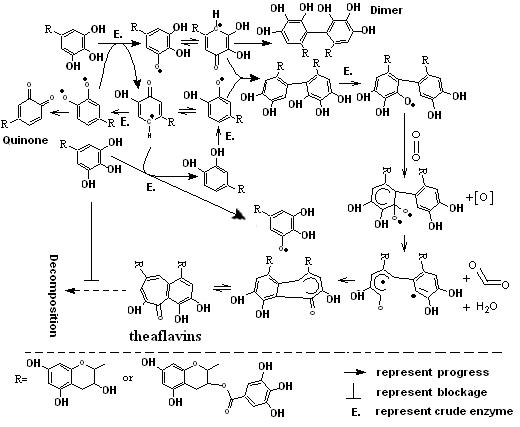
Fig. 5. Proposed possible pathway of theaflavins biosynthesis
Micellar catalysis of surfactants has been widely applied in chemical reactions such as synthesis, substitution, oxidation, and so on[34-37]. Our current data indicated that the concentration of TF-D-G was decreased by the increment of SDS concentration in the same reaction time, which might be due to the low substrate binding affinity of catechins with enzyme in micelle aqueous[37]. But surprisingly, we found the formed micelle system of Sodium dodecylsulfate can help the theaflavins decomposition especially when its concentration over the critical point. So in theory, low-polar ECG entry into the lyophobic micellar core by increasingly aggregated micelle and catalyzed by enzyme to form ECG free radical at the interface[34, 38, 39] and then combined with EGCG free radical which transformed from aqueous phase into the micelle core to form theaflavin in the micelle core (Fig. 6.). Moreover, theaflavins segregated by the lyophobic core of micelle with aqueous enzyme. Thus, higher concentration of SDS can keep more TF-D-G from decomposition, especially in 8 and 16 mmol/ml (Fig. 4.).

Fig. 6. Role of SDS as a catalyst on theaflavins stabilization in aqueous phase
5. Conclusion
The present study indicated the consumed rate of each catechin catalyzed by enzyme independently was taken on similar tendency of each type catechins, along with time progress. The paired catechins of enzymatic reaction with type of simultaneousness or arrangement in order indicated that epigallocatechins such as EGC and EGCG sacrificed themselves to save EC or ECG and generated theaflavins, and prior catalysis with EGC or EGCG was more efficient to form theaflavins in the reaction system than the latter. Moreover, same classified catechins competed with each other for reacting with corresponding catechin. And the reducing agents added can keep the content of catechins especially EGCG in course of enzymatic reaction, but the formation of theaflavin was limited gradually as short as the addition time. Interestingly and most importantly, the SDS as one of the surfactants was found to be positively proportional to the theaflavin generation and can keep it stable in the aqueous solution for the first time.
Acknowledgment
This research was funded by Zhejiang Provincial Natural Science Foundation of China under Grant No. Y3100683. The authors are grateful Shu hua (Department of Tea Science, Zhejiang University) for the harvested fresh tea leaves from the Hua-Jia-Chi tea plant of Zhejiang University.
References
[1] College of Anhui Agronomy, Manufacture of tea. 2nd ed., China Agriculture Press, Beijing, 1979. (In Chinese)
[2] X.C. Wan, Tea biochemistry. 1st ed.,China Agriculture Press, Beijing, 2003. (In Chinese)
[3] E.A.T.H. Roberts, The phenolic substances of manufactured tea (I). J. Sci. Food Agric.8(1957) 72-78.
[4] Dreger H., Lorenz M., Kehrer A., Baumann G., Stangl K. and Stangl V.,Theaflavin-mediated cardioprotection. Exp. Biol. Med.233: 427-433 (2008).
[5] Henning S.M., Aronson W., Niu Y., Conde F., Lee N.H., Seeram N.P., Lee R.P., Lu J., Harris D.M., Moro A., Hong J., Pak-Shan L., Barnard R.J., Ziaee H.G., Csathy G., Go V.L., Wang H. and Heber D.,Tea polyphenols and theaflavins are present in prostate tissue of humans and mice after green and black tea consumption.J. Nutr.136: 1839-1843 (2006).
[6] Kundu T., Dey S., Roy M., Siddiqi M. and Bhattacharya R.K., Induction of apoptosis in human leukemia cells by black tea and its polyphenol theaflavin.Cancer Lett.230: 111-121 (2005).
[7] Lin J.K., Chen P.C., Ho C.T. and Lin-Shiau S.Y.,Inhibition of xanthine oxidase and suppression of intracellular reactive oxygen species in HL-60 cells by theaflavin-3,3'-digallate, (-)-Epigallocatechin-3-gallate, and propyl Gallate. J. Agric. Food Chem.7: 2736-2743 (2000).
[8] Schuck A.G., Ausubel M.B., Zuckerbraun H.L. and Babich H., Theaflavin-3,3'-digallate, a component of black tea: An inducer of oxidative stress and apoptosis. Toxicol. In Vitro 22: 598-609 (2008).
[9] Lu J., Ho C.T., Ghai G. and Chen K.Y., Differential effects of theaflavin monogallates on cell growth, apoptosis, and Cox-2 Gen Expression in cancerous versus normal cells. Cancer Res.60: 6465-6471 (2000).
[10] Lee H.H., Ho C.T. and Lin J.K., Theaflavin-3,3'-digallate and penta-o-galloyl-beta-D-glucose inhibit rat liver microsomal 5alpha-reductase activity and the expression of androgen receptor in LNCaP prostate cancer cells. Carcinogenesis 7: 1109-1118 (2004).
[11] Chung J.Y., Park J.O., Phyu H., Dong Z. and Yang C.S.,Mechanisms of inhibition of the Ras-MAP kinase signaling pathway in 30.7 b Ras 12 cells by tea polyphenols (-)-epigallocatechin-3-gallate and theaflavin-3,3'-digallate. FASEB J.11: 2022-2024 (2001).
[12] Maron D.J., Lu G.P., Cai N.S., Wu Z.G., Li Y.H., Chen H., Zhu J.Q., Jin X.J., Wouters B.C. and Zhao J., Cholesterol-lowering effect of a theaflavin-enriched green tea extract: a randomized controlled trial.Arch. Intern. Med.163: 1448-1453 (2003).
[13] Shen D.Y., Yin J.F., Xu Y.Q., Yuan H.B. and Chen S.Q., Influences of biochemical components in the fresh leaf from different tea cultivars on the ferment effect of black tea. J. Anhui Agri. Sci. 19: 9120-9125 (2009). (In Chinese)
[14] Roberts E.A.H. and Wood D.J., The fermentation process in tea manufacture II. Oxidation of substrates by tea. Biochem. J.47: 175-186 (1950).
[15] Gregory R.P.F. and Bendall D.S., The purification and some properties of the polyphenol oxidase from tea (Camellia sinensis L.).Biochem. J.101: 569-581 (1966).
[16] Halder J., Tamuli P. and Bhaduri A.N., Isolation and characterization of polyphenol oxidase from indian tea leaf (Camellia sinensis ). J. Nutr. Biochem.9: 75-80 (1998).
[17] Ravichandran R. and Parthiban R., Changes in enzyme activities (polyphenol oxidase and phenylalanine ammonia lyase) with type of tea leaf and during black tea manufacture and the effect of enzyme supplementation of dhool on black tea quality.Food Chem.3: 277-281 (1998).
[18] Tu Y.Y., Xu X.Q., Xia H.L. and Watanabe N., Optimization of theaflavin biosynthesis from thea polyphenols using an immobilized enzyme system and response surface methodology. Biotechnol. Lett.27: 269-274 (2005).
[19] Sharma K., Bari S.S. and Singh H.P., Biotransformation of tea catechins into theaflavins with immobilized polyphenol oxidase.J. Mol. Catal. B: Enzy.56: 253-258 (2009).
[20] Bajaj K.L., Anan T., Tsushida T. and Ikegaya K.,Effects of (-)-epicatechin on oxidation of theaflavins by polyphenol oxidase from tea leaves.Agric. Biol. Chem.7: 1767-1772 (1987).
[21] Tanaka T., Mine C., Inoue K., Matsuda M. and Kouno I., Synthesis of theaflavin from epicatecin and epigallocatechin by plant homogenates and role of epicatechin quinone in the synthesis and degradation of theaflavin. J. Agric. Food Chem.7: 2142-2148 (2002).
[22] Owuor P.O. and Obanda M., The use of green tea (Camellia sinensis) leaf flavan-3-ol composition in predicting plain black tea quality potential. Food Chem.100: 873-884 (2007).
[23] Dong H.Z., Zeng S.F., Gao D.F., Yang S.R., Zhang Y.J. and Jiang H.J., Primary analysis of chemical components of Black tea from Yunnan province. Tea Sci. Technol.2: 14-17 (2009). (In Chinese)
[24] Leung L.K., Su Y., Chen R., Zhang Z., Huang Y. and Chen Z.Y., Theaflavins in black tea and catechins in green tea are equally effective antioxidants.J. Nutr.131: 2248-2251 (2001).
[25] Wang K.B., Liu Z.H., Huang J.A., Dong X., Song L., Pan Y. and liu F., Preparative isolation and purification of theaflavins and catechins by high-speed countercurrent chromatography.J. Chromatogra. B867: 282-286 (2008).
[26] Li Y., Shibahara A., Matsuo Y., Tanaka T. and Kouno I., Reaction of the black tea pigment theaflavin during enzymatic oxidation of tea Catechins. J. Nat. Prod.73: 33-39 (2010).
[27] Cloughley J.B. and Ellis R.T., The effect of pH modification during fermentation on the quality parameters of central African black teas. J. Sci. Food Agric.31: 924-934 (1980).
[28] Subramanian N., Venkatesh P., Ganguli S. and Sinkar V.P., Role of Polyphenol oxidase and peroxidase in the generation of black tea theaflavins.J. Agric. Food Chem.7: 2571-2578 (1999).
[29] Sellés-Marchart S., Casado-Vela J. and Bru-Martínez R., Effect of detergents, trypsin and unsaturated fatty acids on latent loquat fruit polyphenol oxidase: Basis for the enzyme’s activity regulation. Arch. Biochem. Biophys.464: 295-305 (2007).
[30] T. Tanaka, Y. Miyata, K. Tamaya, R. Kusano, Y. Matsuo, S. Tamaru, K. Tanaka, T. Matsui, M. Maeda and I. Kouno, Increase of theaflavin gallates and thearubigins by acceleration of catechin oxidation in a new fermented tea product obtained by the tea-rolling processing of loquat (Eriobotrya japonica) and green tea leaves.J. Agric. Food Chem.57 (2009)5816-22.
[31] H.S. Joglekar, S.D. Samant and J.B. Joshi, Kinetics of wet air oxidation of phenol and substituted phenols. Wat. Res.2 (1991)135-145.
[32] W. Li, Q. Guo, B.L. Zhao, S.R. Shen, J.W. Hou and W.J. Xin, ESR spectroscopy structural analysis of the semiquinone free radicals of green tea polyphenols and their components. Chinese J. Mag. Reson. 6 (1998)507-513. (in Chinese)
[33] J. Zhu, X.G. Meng, R.R. Zang, M.Z. Chen, C.W. Hu, and X.C. Zeng, Kinetic study of oxidative coupling reaction of 2,6-dimethylphenol catalyzed by copper(II) complexes.Chinese J. Catal.4 (2005)317-322. (In Chinese)
[34] B. Boyer, A. Hambardzoumian, J.P. Roque and N. Beylerian,Reaction in biphasic water/organic solvent system in the presence of surfactant: inverse phase transfer catalysis versus interfacial catalysis. Tetrahedron 56 (2000)303-307.
[35] L.J. Mathias and R.A Vaidya., Inverse phase transfer catalysis. First report of a new class of interfacial reactions. J. Am. Chem. Soc. 108 (1986)1093-1095.
[36] J.Z. Zhou and Z. Wen, Progress of micellar catalysis and its mimic enzyme. Chem. Bull. 5 (1987)21-26. (in Chinese)
[37] Y.L. Zhou, X.Q Fu., L Ma., and J.D. Wang, Effect of microenvironment of substrates on the catalysis of horseradish peroxidase in aqueous micelle system. Chem. J. Chinese U.12 (1996) 1927-1929. (In Chinese)
[38] J.B.F.N. Engberts, Catalysis by surfactant aggregates in aqueous solutions. Pure Appl. Chem.11 (1992)1653-1660.
[39] P.V. Shinde, A.H. Kategaonkar, B.B. Shingate and M.S. Shingare, Surfactant catalyzed convenient and greener synthesis of tetrahydrobenzo [a] xanthenes-11-ones at ambient temperature. Beilstein J. Org. Chem. 7 (2011)53-58.
https://blog.sciencenet.cn/blog-71940-694191.html
下一篇:第一次申请省科学基金与结题小结