博文
科学遐想-神秘的缠绕(7--12)
||
七、神秘的缠绕-RNA:DNA纠缠
我在英国爱丁堡大学发现,人类基因组中分布的10类三核苷酸重复序列的转录有些会受到严重影响,比如GAATTC序列(DNA repair 2009年),而其他与疾病有关的正常人体内三核苷酸重复CTG/CAG, CGG/CCG 并不明显影响基因转录,只是频繁地发生 “Slippage”(DNA 复制过程中模板链和新合成的互补链频繁地以“CAG”,“CTG”,“CGG” 和“CCG”为单位发生“Indel”(插入-缺失)改变。而达到一个长度“阈值”会突然扩增,变得更长。
受那些影响基因转录的三核苷酸重复的启发,2006年,我第一次提出与疾病相关的“CAG/CTG”,“CGG/CCG”等在特定长度时应该也会影响基因转录,比如当这些序列形成其他非B型构象时,对RNA转录的影响有可能使得RNA 无法与DNA模板分离,形成RNA:DNA 杂交体(R Loop)。而这种错误的“缠绕” 需要处理,造成非模板DNA链断裂,而与模板链结合的RNA被“误作”引物被用于引导DNA复制。在这个过程中,拓扑缠绕起着关键的作用,而非模板链的断裂势必会“缓解” R 环部位的“拓扑学缠绕”(情同DNA拓扑异构酶I的 切断)。
2015年,这个假说被西班牙科学家利用其他重复DNA序列给予了证实。他们发现,在重复序列中出现的R环中的RNA 半分子确实被用于引导DNA 复制。
随后,有关研究的发生了转向,2022年人们开始围绕我最先的推想(我最先提出的工作机制)探索疾病患者体内这些疾病相关的三核苷酸重复序列异常扩增的可能机制。
这就是当前相关疾病病理机制研究的大致情形。
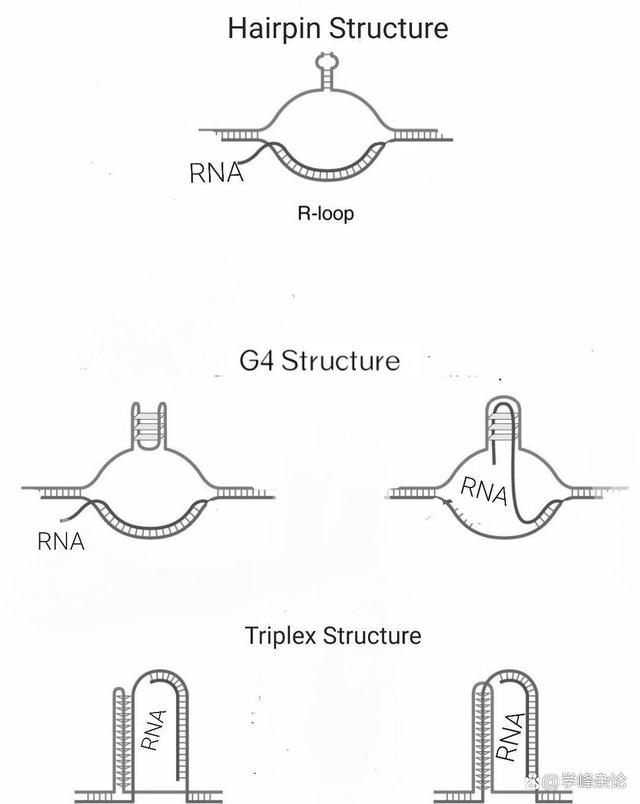 RNA 转录与非B-DNA,2022
RNA 转录与非B-DNA,2022
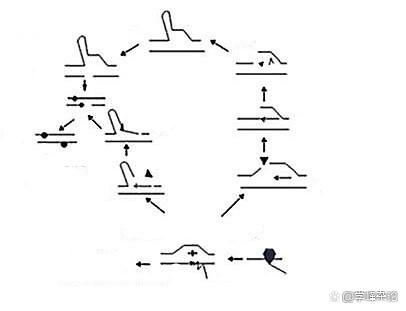 我最先提出的工作机制 2006
我最先提出的工作机制 2006
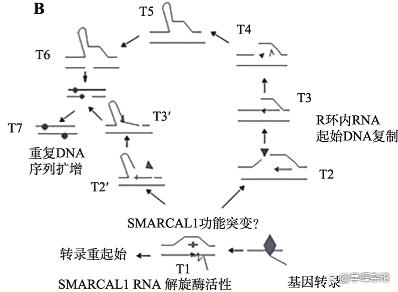 注解
注解
这个机制的重要意义在于它和50种目前尚无计可施的人类神经-肌肉系统退行性疾病的发生密切关系。
八、神秘的缠绕-引物酶来自拓扑异构酶
众所周知,作为遗传信息载体的DNA分子需要在细胞繁殖过程复制一次,以倍增“基因组”。
这种基因组DNA的复制所需要的DNA聚合酶只能催化新生DNA链沿着5'-3'方向进行,而对于以5'-3'和3'-5'反向互补DNA链而言,位于DNA复制叉内的DNA聚合酶也只能以“反向”同时复制双链DNA分子,而这种“反向”复制需要一条链“翻转”与其互补链“同向”,而复制叉的这种布局使得翻转的那条链的复制不能“连续”,需要反复合成RNA 引物,这需要其复制酶(引物酶,Primase)一直“伴随”复制叉。
引物酶与DNA模板结合
引物酶与DNA结合力较弱,因为引物酶与其他DNA依赖的RNA聚合酶在结构上明显不同,表明它们之间没有“共同”起源(这一点很重要,因为在没有发现引物酶之前,人们曾一度认为DNA复制所需要的RNA引物可以由负责基因转录的DNA 依赖的RNA聚合酶提供,这在一些细菌的质粒DNA 和线粒体DNA复制时已经被确认)。
引物酶所含有保守的“Toprim”结构域作为催化中心的分子结构表明它自身与拓扑异构酶IA和II具有共同的起源。这或许可以帮助我们理解它不能与DNA模板紧密结合的原因。
即使如此,为了能“不离不弃”地追随DNA复制叉,引物酶“进化”出以下“能力”:
1、与负责DNA复制的解旋酶在蛋白水平形成复合体,如大肠杆菌的DnaB-DnaG, 其中,DnaB 是具有六元环结构的DNA解旋酶,以5'-3'方向沿着DNA复制叉中的“滞后链”(lagging strand template)向着双链区方向“滑动”,由于这个DnaB 六元环“套住”了DNA 链,不易滑落,所以DnaG 引物酶与之结合可以解决其与DNA 模板结合不牢固的问题;
2、在基因水平与解旋酶编码区“组装”
以大肠杆菌T7噬菌体的“引物酶-解旋酶”为例,C端的“解旋酶结构域”以六元环结构与DNA模板结合,负责解开复制所需要的单链模板,而N端的引物酶结构域则负责合成“引物”。引物酶识别序列5'-GGGTC-3'、5'-TGGTC-2'和5'-GTGTC-3'。并在相应序列处合成功核糖核苷酸引物pppACCC、pppACCA和pppACAC,这些引物酶识别位点包含基本识别序列5'-GTC-3',其中,识别3'-胞苷为引物酶识别的必需碱基,但这个碱基不复制到引物中。在序列5'-GTC-3',引物酶催化核糖核苷酸pppAC的合成,但只有四个核糖核苷酸的长度才能作为T7 DNA聚合酶的有效引物。
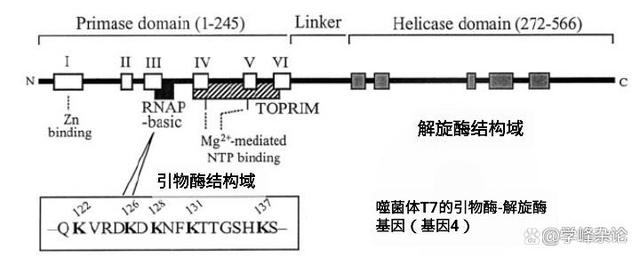 T7噬菌体的基因4
T7噬菌体的基因4
3、与DNA复制叉中其他蛋白质的广泛结合能力
除了在蛋白水平和基因水平与DNA复制解旋酶“绑定”和“整合”之外,真核物种中的RNA 引物酶采取与DNA聚合酶alpha 形成复合体
 参与真核基因组复制的聚合酶-引物酶
参与真核基因组复制的聚合酶-引物酶
4、与其他“复制”蛋白结合。
九、神秘的缠绕-Toprim域工具
Toprim(拓扑异构酶原酶)结构域是一个由约100个氨基酸组成的结构保守结构域,见于细菌DnaG型引物酶(Primase)、来自细菌和古菌的小引物酶样蛋白(small primase- like)、IA型和II型拓扑异构酶、OLD家族的细菌和古细菌核酸酶以及RecR/M家族的细菌DNA修复蛋白中。
Toprim结构域可以单独存在于一种蛋白质中,也可以与几个其他的结构域组合,如与ASM结构域、超家族2解旋酶结构域、超级家族3解旋酶结构域、DnaB作用结构域、C4“小指”结构域、CHC2锌指结构域、HSP90-螺旋酶-组氨酸激酶超家族的ATPase结构域、S5结构域、SET结构域,ABC转运蛋白/SMC超家族的螺旋-发夹-螺旋(Helix-hairpin-Helix)DNA结合结构域、动员(mobilization, MOB)结构域或ABC转运蛋白-SMC超家族(ABC transporter/SMC superfamily)ATPase结构域共存于一种蛋白质。
Toprim结构域是拓扑异构酶能切断DNA单链或/双链后可以重新将断裂连接的“催化”结构域。是否在其他蛋白质内也发挥类似的DNA链断裂和重新连接或可能发挥的作用值得关注。
Toprim结构域有两个保守基序(motif),其中一个中心含有一个保守的谷氨酸,另一个含有两个保守天冬氨酸(DxD)。这两个基序之前都有保守的疏水区域,预计将形成β-折叠片(beta strand)。谷氨酸残基可能参与催化作用,而"DxD"基序负责与二价镁离子通过配位键形成“Toprim”酶活性所中心。
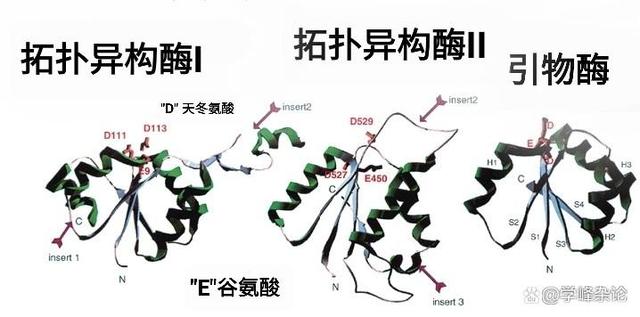 Toprim,D E 分置于两个基序
Toprim,D E 分置于两个基序
Toprim结构域具有紧凑的α/β折叠,具有四个保守的beta-折叠片层和三个alpha-螺旋;除了第二个alpha-螺旋和C-末端的beta-折叠片层之外,其余的二级结构在Toprim 结构域中元件中的位置高度保守。Toprim结构域包含三个可容纳可变大小“插入”区域,这已经在拓扑异构酶中得到了证实。
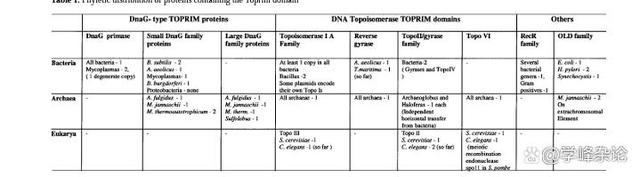 三界生物含TOPRIM 蛋白
三界生物含TOPRIM 蛋白
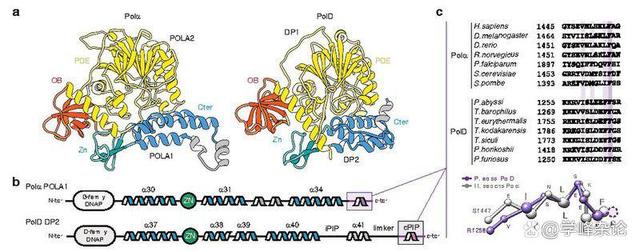 引物酶-真核生物
引物酶-真核生物
十、神秘的缠绕-转录延伸
DNA负载的“信息” 可以通过“转录” 转移至RNA媒介分子,包括用于编码多肽的mRNA, 用于搬运氨基酸的tRNA, 作为“催化剂”的 rRNA, 以及非编码多肽信息的其他非编码RNA (noncoding RNA, ncRNA) , 如siRNA 等等。
转录需要面对DNA 分子缠绕,由于DNA双螺旋是右手负超螺旋,因此,RNA聚合酶前行方需要利用引入左手正超螺旋,而在RNA聚合酶后方则需要负超螺旋把打开的DNA双链重新“旋紧”。这种缠绕引起的DNA双链的“旋转”会使整个RNA聚合酶分子在DNA模板上旋转。同样,所转录出的 RNA分子也会随之“缠绕”,并有可能重新与其DNA模板形成RNA:DNA杂交体(RNA环,R-LOOP)。
 RNA转录过程发生的缠绕
RNA转录过程发生的缠绕
 R-LOOP 与DNA 分子缠绕
R-LOOP 与DNA 分子缠绕
以上的情形常用于描述那些不与蛋白质结合的DNA分子的“转录”,如原核生物基因组DNA分子的转录。众所周知,真核生物细胞内的DNA分子经常与组蛋白八聚体“缠绕”呈核小体形式,由于这些核小体结构被用于“抵消”DNAZ分子的负超螺旋(以左手缠绕146bp,1.75或1.65圈,这近乎两圈的缠绕分别缠在核心组蛋白八聚体的H2A-H2B,H3-H4半分子上),显然,基因转录时“不能”把被转录区段的核心组蛋白八聚体从染色质上全部剥离(通过对组蛋白N端无序肽链中的碱性氨基酸修饰乙酰基团,来自乙酰辅酶A, 由三梭算循环产生,改变组蛋白与DNA磷酸骨架的稳定状态松开缠绕DNA),那样会在本该转录的染色质区段形成"precatenanes” 。
真核生物染色质转录过程必须面对组蛋白八聚体缠绕问题
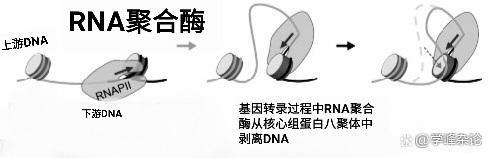 缠绕在组蛋白八聚体上的DNA剥离
缠绕在组蛋白八聚体上的DNA剥离
与组蛋白八聚体缠绕的DNA通过被剥离,慢慢以环的形式“抽出”用于转录模板,而非完全去除组蛋白八聚体。
以下是真核染色质转录的可能过程,其中“缠绕”的解除伴随着组蛋白八聚体的“半分子”拆开,拆开后的DNA分子依然和八聚体半分子“缠绕”着。
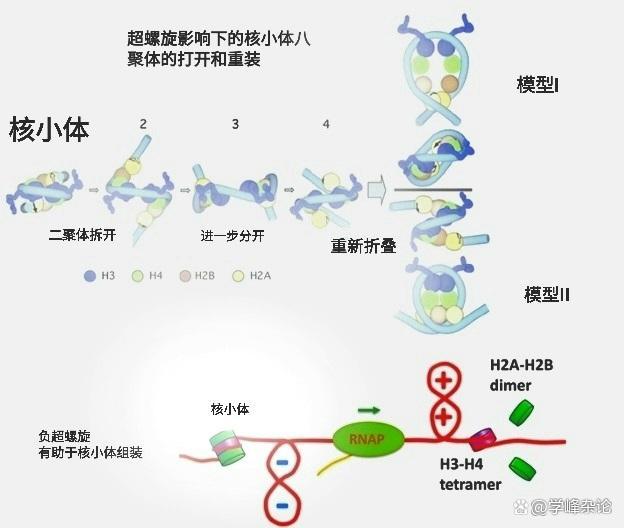 真核染色质的转录
真核染色质的转录
DNA分子转录“泡儿”前方的正超螺旋有助于上述组蛋白八聚体半分子的拆开,而其后方的负超螺旋则有助于组蛋白八聚体的重新形成。
十一、神秘的缠绕-跨越核小体
染色质由DNA缠绕核心组蛋白八聚体形成的核小体形成“串珠”,其中组蛋白尾巴上的碱性氨基酸残基通过正电荷与磷酸骨架上的负电荷发生静电相互作用。这会阻碍基因转录过程中RNA聚合酶的通过。
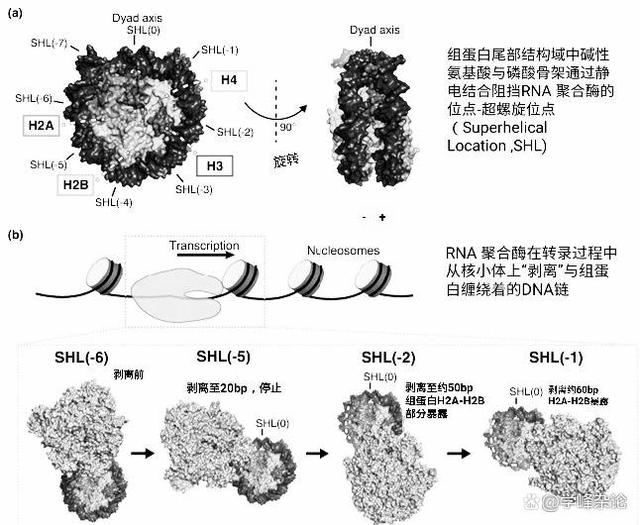 从二元轴开始,每10个碱基对为一个
从二元轴开始,每10个碱基对为一个
从二元轴开始,每10个碱基对定名为一个超螺旋位点(SHL),1.75圈约146个碱基对,左右约可分为7个SHL位点(2×70bp=140bp)。
RNA 聚合酶与组蛋白伴侣和转录延伸因子一道从核小体中“剥离” DNA。
DNA聚合酶自身具有“剥离”DNA的能力
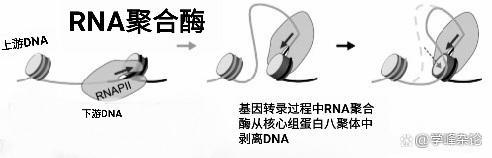 聚合酶自身有剥离DNA能力
聚合酶自身有剥离DNA能力
尽管RNA聚合酶自身拥有从核心组蛋白八聚体上“剥离”缠绕的DNA进行基因转录,但为了提高效率,不致于造成RNA聚合酶停止甚或从模板上脱落,组蛋白伴侣和转录延伸因子会“协助”RNA聚合酶。
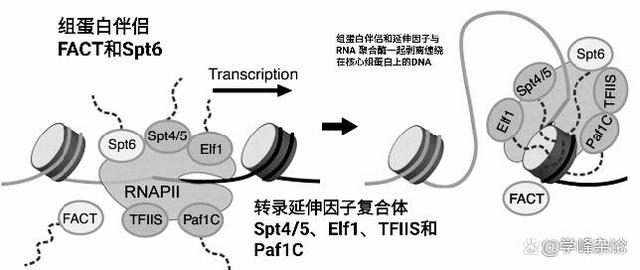 从核小体中剥离DNA
从核小体中剥离DNA
重塑因子负责调整“变动”后的核小体分布
 转录过程中的核小体重塑
转录过程中的核小体重塑
总结
基因转录过程中需要“跨越”核小体“障碍”。RNA聚合酶、组蛋白伴侣、转录延伸因子一道把缠绕在核心组蛋白八聚体上的DNA剥离,造成的核小体“疏远”和“拥挤”由核小体重塑因子负责再调整。
十二、神秘的缠绕-是什么吸引了复制起点蛋白
真核生物基因组中DNA复制起点是如何“确定”的?这是个一直都没有明确“答案”的问题。比如,人的基因组的DNA复制通常在数万个位点(30000-50000处)开始复制,这些位点需要在细胞周期的G1时相结合ORC (Origin Recognition Complex)蛋白,之后,利用Cdc6-Ctd1 装载MCM2-7(解旋酶6聚体,相当于大肠杆菌细胞中的DnaB)。最近自然杂志发表了2篇文章(2022),一篇发现真核生物基因组TAD结构域及子TAD (subTAD与复制起始位点选择的可能相关性,这个工作试图依据结构和分子特征对TAD和子TAD进行分层。在全局和局部微扰的情况下,揭示了在S期启动DNA复制情况下,G1/pre-S中由cohesin (粘蛋白)介导的环形成拥有“局限”基因组DNA 复制起始的功能,主要是发现“局限”了MCM在基因组范围的“分布”/“富集”。另一篇文章则明确地表明,MCM可以“阻碍” cohesin、CTCF等催化的DNA looping。与此有关的一个工作表明,MCM可以“随机”与DNA分子结合,MCM过表达并不会影响复制的整个过程,包括在其在细胞周期S时相的"firing"等,相反,低水平的MCM确实会影响基因组复制。
综合三方工作,似乎支持如下情形,首先未被磷酸化修饰激活的MCM可以随机与DNA分子结合,这种结合会影响cohesin 和CTCF介导的染色质成环(looping out),因此另一方观察到的MCM与TAD域和子TAD域的差异性分布也有可能是因为受到Cohesin 和CTCF等的影响,不再能在染色质中“自由”扩散有关。
有研究工作表明,在真核基因组DNA 复制起点确定过程中,只有ORC蛋白能结合的DNA 位点才是真正的潜在的复制起始位点,而只有与ORC结合的MCM才有可能真正在细胞周期S时相被磷酸化修饰得到“激活”,参与DNA复制。
最近与ORC蛋白与DNA位点结合的研究发现以下现象:
1、 在果蝇和小鼠等后生生物基因组中,绝大多数的CpG岛(CGI,CpG Islands)附近含有DNA复制起始位点Ori。尽管果蝇中几乎不存在甲基化,但大多数CpG岛(CGI)都含有ORI,这表明这种表观遗传学标记对于定义激活起源并不重要。DNA合成起始于CGI的边界,导致新生链呈明显的双峰分布,暗示了双重起始事件。
2、 富含GC的重复序列元件可用于后生生物基因组复制起点(Oris)的良好预测因子,表明后生生物基因组DNA复制起点也体现部分一致的碱基排列特征。
3、 通过标记新合成的DNA新生链发现,Ori处含有与新生链峰一致的独特核苷酸偏好,其特征是Ori位点5'和3'的G/T和C/A过度性存在。
4、 全基因组分析与DNA梳理分析相结合发现基因组中含有大量的可作为ORI的组织结构。但是,这些ORI的激活并不随机发生。它们被组织成特定位点,灵活使用ORI,可推测,这些Ori确定了一个复制子(Replicon),在每个复制子中激活(Firing)一个Ori。这样的组织使得每个复制子具有位点特异性和Ori firing的灵活性,可能与环境和细胞命运不同适应有关。
5、 小鼠胚胎干细胞中85%的复制起始位点与转录单位有关。近一半的已鉴定ORI图谱位于启动子区域,有趣的是,ORI密度与启动子密度密切相关,反映了小鼠基因组中复制和转录的协调组织。
6、 对ORI活性的详细分析表明,CpG岛启动子-ORI是胚胎干细胞(ES)中最有效的ORI,并且ORI特异性和点火(Firing)效率在不同细胞类型中都保持不变。而且启动子-ORI处复制起始位点的分布与转录起始位点(TSS)的分布完全平行,表明驱动复制和转录的调控区域共同进化。
7、 RNA:DNA杂交体的形成区和DNA复制起始位点的相关性。R-环与ORC共同定位在同一个CpG岛区域。这些有效的DNA复制起始位点的很大一部分正好位于G4(四链体)基序密度最高的位置。这种情景暗示人类细胞内基因转录和DNA复制之间的联系,并表明CpG岛启动子-DNA复制起始位点处形成R-环诱发的DNA复制失调可能与癌症细胞出现的DNA复制异常和基因组完整性缺失有关。
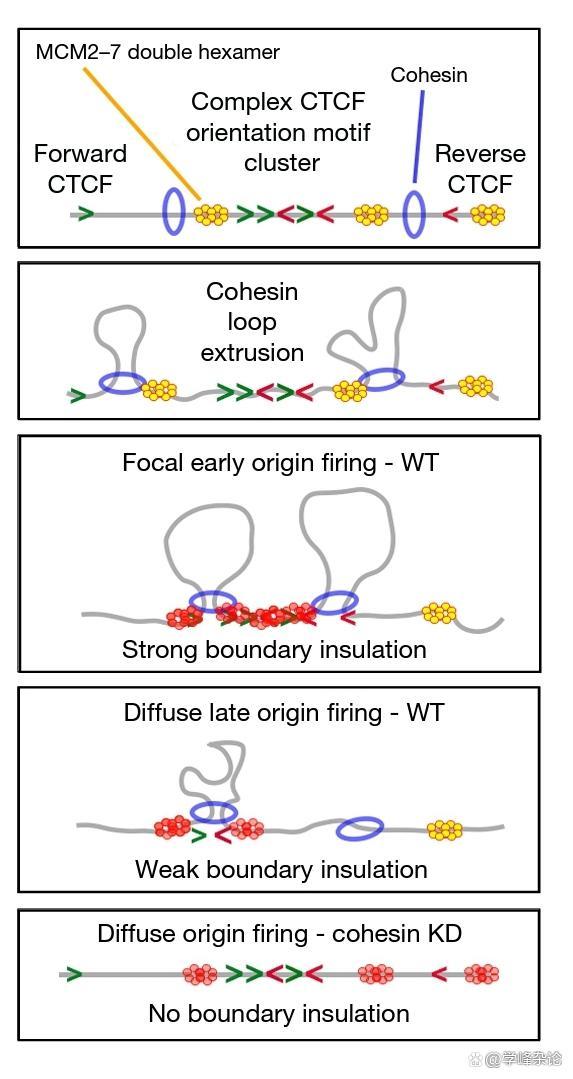 TAD限制MCM的富集
TAD限制MCM的富集
https://blog.sciencenet.cn/blog-218980-1352861.html
上一篇:科学遐想-神秘的缠绕(1-6)
下一篇:分子医学基础-不完美的设计-酷蛋白与肿瘤(13-16)