博文
猴痘病毒(1):起源、传播和进化
||
猴痘病毒起源、传播和进化
在长期的自然进化中,病毒作为一个存在实体,与宿主和环境持续相互作用,在一定的时间内形成了相对稳定的生态位。随着人类活动以及由此引起的气候和环境的改变,病毒与宿主的相互作用也在发生改变,之前相对稳定的生态位会被打破,进而对病毒的进化产生重要影响。
半个多世纪以来,猴痘病毒的进化一直伴随着病毒传播模式的改变。自1970年代首次报道人感染猴痘病毒以来,猴痘主要在中非(刚果盆地)和西非地区以人兽共患模式呈地方性流行。进入新世纪以来,猴痘的暴发呈上升趋势,并出现人际间传播,但主要还是发生在上述地方性流行地区。2005-2018年,刚果每年报道1000多例确诊或/和疑似病例。2003年,美国确诊了47例猴痘病例,大部分病例接触过发病的草原犬鼠,但没有发现人传人病例;流行病学调查表明后者都曾经暴露过从西非加纳进口到美国的一些啮齿类动物。这是发生在非洲以外人感染猴痘的首次报道]。2017-2018年,尼日利亚发生了120多例人感染猴痘的暴发疫情。2018-2021年,非洲之外的一些国家(包括美国、英国、新加坡和以色列等)也发现了与尼日利亚疫情相关的输入病例。与之前的暴发相比,2022年5月以来猴痘病毒的传播表现出一些新的特征,传播的地理范围持续扩大,人传人的特征更加明显,病毒的传播力更强,这引起了人们对猴痘病毒的变异及其进化的广泛关注。
研究发现,所有正痘病毒属成员都是由一个共同的祖先病毒进化而来。2010年以来,在美洲的人和灵长类中发现了2种不同于旧世界(欧亚大陆和非洲)的正痘病毒,表明正痘病毒地理分布和进化的复杂性超出人们之前的认识。最近Babkin等对正痘病毒基因组中央区约102kb长的保守序列进行分子进化分析,结果表明约在40000年前美洲大陆和旧世界的正痘病毒出现分化,而大约在3500年前猴痘病毒从旧世界的其它正痘病毒中逐渐分化出来。
传统上,根据病毒基因序列,猴痘病毒可以分为两个具有明显地理特征的进化分支,分别是中非(刚果盆地)分支和西非分支,分支间的核苷酸同源性约为95%,分支内同源性高达99%。分子进化分析认为,西非分支是大约在600年前分化出来的。这两种不同分支的病毒在致病性和传播力都不尽相同,其中西非分支的症状较轻,病死率大约是3.6%(95% CI 1.7-6.8%);而中非分支引起的症状较重,病死率可达10.6%(95% CI 8.4-13.3%)。
最近WHO推荐了猴痘病毒一种新的进化分类法,将病毒分为I(刚果盆地或中非分支)和II(西非分支)两大分支,其中分支II包括IIa和IIb两个亚分支。IIb是从IIa进化而来,主要由1970年以来感染人的毒株构成。在IIb中,2017-2022年以来的毒株表现为持续人际间传播特点,故称为人猴痘病毒分支1(hMPXV1),hMPXV1又进一步分分为A、A.1、A.2、B1等谱系。A.2谱系包括2017-2018年尼日利亚暴发毒株,A.1谱系包括2018-2021年新加坡、英国、美国和以色列输入性毒株。2022年疫情的毒株聚集成B1谱系,由A.1谱系分化出来,表明这些毒株有共同单一的遗传起源。
猴痘病毒分支I和II之间基因组变异最大的区域主要集中在ITRs的末端区域,而ITRs主要编码宿主应答调节子(HRM)蛋白的基因。与分支I相比,分支II的ITRs缺失了编码猴痘病毒补体酶抑制子蛋白(MOPICE)基因,这可能是分支II病毒致病性减弱的一个原因,但对这一推论仍存在争议。分支I和分支II的致病性差异,可能并非由单个基因决定,而是受病毒多个基因的影响,包括D10L、B10R、B14R和B19R等开放阅读框架。
与2017-2021流行的猴痘相比,2022疫情症状更轻,对人体的适应力和人际间传播力更强。人们据此推测,B1谱系病毒被发现前,可能已经在世界某个地区的人群中隐匿流行了较长一段时间。这个起源地可能是猴痘传统的地方性流行区(西非和中非),也可能是非洲之外的非地方性流行区。在隐匿的流行过程中,病毒持续进化,渐次增强对人体的适应力。最近的一些研究支持了这个假说。Isidro等利用鸟枪法宏基因组技术,分析了2022年5月欧洲15株早期病例的毒株,系统进化分析表明病毒与2017-2019年尼日利亚暴发流行的毒株有亲缘关系,是后者持续进化的产物。基因组深度分析发现,与2018-2019年的病毒相比,2022年IIb-B1谱系病毒的基因组中出现了约50个单核苷酸多态性(SNPs)的遗传差异。据估算,正痘病毒的核苷酸进化速率一般为每年每个基因组发生1-2个核苷酸替代(2 × 10−6 –1 × 10−5 核苷酸替代/位点/年)。但IIb-B1谱系病毒的的核苷酸替代率增加了6-12倍,提示IIb-B1谱系病毒的进化速率明显在加快。
进一步分析发现,B1谱系病毒有46个SNPs表现出突变偏倚,其中26个和15个分别替换为GA > AA和TC > TT。病毒基因组的这种突变偏倚和A:T碱基丰富度的现象,提示了在病毒进化过程中可能存在一种非随机驱动因素。研究者据此推测,宿主表达的载脂蛋白B mRNA编辑催化多肽样3蛋白(APOBEC3)可能是IIb-B1谱系病毒进化的重要驱动因素[19]。APOBEC3被认为是人和哺乳动物抗病毒先天免疫功能的一个构件,病毒感染会引发APOBEC3表达上调,通过脱氨酶作用和脱氨酶非依赖性机制来介导病毒的基因组突变。APOBEC3驱动的突变一方面可能会抑制病毒的复制,另一方面也可会使病毒产生突变体。APOBEC3的这种功能也出现在一些其他病毒的感染过程中,如HIV、乙型肝炎病毒(HBV)、人类乳头状瘤病毒(HPV)、单纯疱疹病毒(HSV)和EB病毒(EBV)等。J.P.Thornhill等人的研究也发现,528例猴痘病例中,218例(41%)也是APOBEC3表达水平较高的HIV患者,猴痘病毒感染HIV患者可能更有利于产生上述突变偏倚。 总之,人体APOBEC3蛋白可能是引起B1谱系病毒进化加速的一个重要因素,使突变的病毒增强了对人体的适应力和传播力,同时降低了致病力。未来的工作需要进一步通过功能性研究等,来对上述推论进行实证回答。
此外,研究也发现,与2018-2019年流行的病毒相比,B1谱系病毒的表面糖蛋白B21发生了3个氨基酸的突变(D209N、P722S和M1741I)。B21是一个重要的体液免疫靶蛋白,其突变有利于病毒免疫逃避。因此,猴痘病毒在传播过程中的抗原漂移也是一个值得关注的现象。
正痘病毒的进化方式在基因组层面包括点突变、基因缺失、基因重组和基因增倍等。猴痘病毒是一种DNA病毒,病毒在复制过程中的DNA聚合酶有3`-5`外切酶校对活性,因此理论上点突变的频率远低于RNA病毒。宿主APOBEC3蛋白驱动的适应性点突变,是2022年疫情B1谱系病毒的主要进化方式。 B1谱系病毒在全球的时空传播过程中,一直伴随着病毒点突变引起的微进化( microevolution),并逐渐分化出不同的小分支(如B.1.1, B.1.2, B.1.3, B.1.4, B.1.5, B.1.6, B.1.7, and B.1.8)。Gigante等发现2022年美国流行的206个毒株中,除了点突变外,有7个毒株出现基因缺失(跨度2.3kb-15kb)或者基因插入-缺失现象,主要分布在病毒基因组的末端,推测这可能会影响病毒的宿主改变和毒力[25]。可以预见,病毒在未来的传播进化过程中,基因组可能会采取的不同进化方式,对病毒的表型和生物学特性都可能产生不同程度的影响。在目前全球猴痘流行的背景下,需要持续监测的猴痘病毒的分子进化及其表型改变,及时调整防控策略和防治手段。
参考文献(略)

图1.猴痘病毒的流行简史
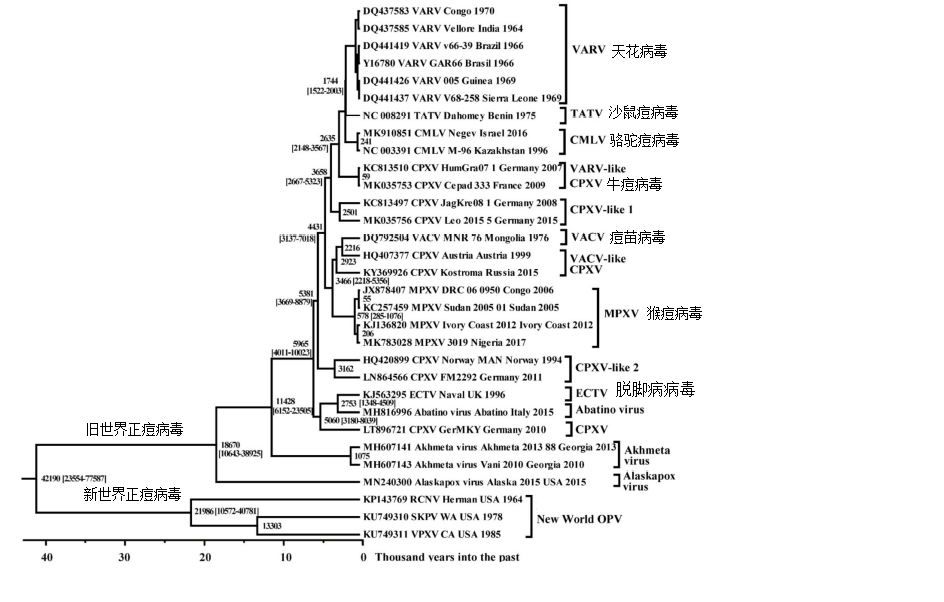
图2.正痘病毒进化树(Babkin等,Viruses,2022)。
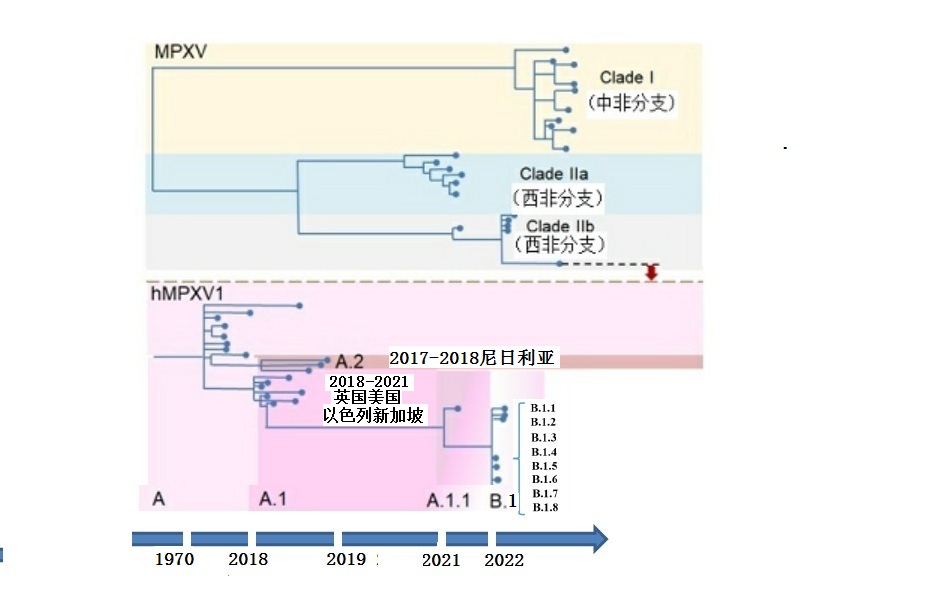
图3.猴痘病毒的分子进化(Chakraborty等, Geroscience ,2022,略加修改)
https://blog.sciencenet.cn/blog-3245311-1386555.html
上一篇:病毒群像(1)--猴痘病毒
下一篇:猴痘病毒(2):检测技术与研究展望