博文
Molecular Plant:青蒿等位基因染色体水平基因组组装揭示了ADS扩增与青蒿素产量之间的相关性
||
发表时间: 2022-08-01
杂志:Molecular Plant
第一作者:廖保生,徐江,郭帅,沈晓凤
通讯作者:陈士林,徐江、张伯礼
机构:中国中医科学院中药研究所,天津中医药大学,中国中医科学院青蒿素研究中心
2022-09-15 16:58
复制
青蒿等位基因染色体水平基因组组装揭示了ADS扩增与青蒿素产量之间的相关性
摘要
青蒿是全球常用抗疟疾药物青蒿素的主要来源。在这里,我们展示了两种青蒿素含量不同的青蒿的染色体水平单倍体图谱,以探索基因组组织与青蒿素生产之间的关系。高保真测序、光学作图和染色质构象捕获测序用于组装异质和重复基因组并解析青蒿的单倍型。每个单倍型基因组注释了大约 50,000 个基因,并且首次在该物种中发现了大约 5812 万年前发生的三倍体事件。在单倍型之间的成对比较期间,在 1.5-Gb 基因组中鉴定出总共 3,903,467–5,193,414 个变体(SNP、插入缺失和结构变体),与该物种的高杂合性一致。基因组分析揭示了青蒿素浓度和紫穗槐-4,11-二烯合酶基因(ADS)拷贝数之间的相关性。通过对具有不同青蒿素含量的 36 个青蒿样品进行重新测序,进一步证实了这种相关性。转录本的CCSa有助于检测旁系同源物表达。总的来说,我们的研究为两种青蒿提供了染色体水平的等位基因感知基因组组装,并为青蒿素的生物合成及其调控提供了新的见解,这将有助于在全球范围内战胜疟疾。
关键词
#青蒿素 #青蒿 #拷贝数变异 a #紫穗槐-4,11-二烯合酶 #基因组 #单倍型
问题
1.用了哪些实验材料及原因?
• 两个黄花蒿(Artemisia annua)品系:高青蒿素系HAN1,低青蒿素系LQ-9(用于基因组组装,分别进行PE150,PacbioHIFI,Hi-C);
• 36 个品种(用于重测序)
• 两个株系同一时期的侧根(根)、二级分枝(茎)、二级分枝上的叶(叶)、花(花)(转录组测序-PE350,其中叶、茎和根使用PacBio Sequel I平台进行三代转录组测序)
2.实验方法及用途?
• Pacbio HIFI
• Hi-C
• PE150 (270 bp and 500 bp,350bp文库)
• DLE-labeling optical maps
• RNA-seq(350bp文库,cDNA SMRTbell文库)
• WGCNA,
3.主要的结果有哪些?
• 获得两个高杂合黄花蒿单体型基因组
• 黄花蒿进化历程推断
• 黄花蒿具有丰富的遗传多样性
• 获全基因组黄花蒿基因表达情况和差异情况,得到ABP通路相关基因共表达网络
• 对ABP基因进行全基因组范围扫描和结构分析,确定ABP途径中ADS基因的拷贝数变异与青蒿素含量具有强相关性。
• 采用液相色谱-质谱联用法测定了黄花蒿GSTs中青蒿素的含量,推测ABP活跃的非GST细胞仍能生产青蒿素。
• ADS基因是进行优良黄花蒿选种的最佳遗传标记
4.创新点在哪?
• 技术新,第一个使用Hi-Fi + DLE + Hi-C进行非模式物种测序和分型的研究;
• 聚焦与ABP通路中,对已知ABP通路中的基因进行了全基因组扫描和分析,确定了ABP基因的表达模式和位置结构特征。
• 使用PacBio Iso-Seq对黄花蒿进行三代转录组测序,并聚焦与ABP通路。
• 结合变异和表达分析,确定了ADS基因的拷贝数变异与青蒿素含量的强相关性,可以将其作为黄花蒿优良选种的遗传标记
• 首次采用液相色谱-质谱联用法测定了黄花蒿GSTs中青蒿素的含量,推测ABP活跃的非GST细胞仍能生产青蒿素。
5.研究思路?
• 首先进行两个单倍体基因组组装;
• 接着对黄花蒿进行比较基因组分析,推断基因组进化情况
• 基因组内变异检测,摸清基因组整体变异情况,观察是否有特定基因受到选择。
• -整体基因组中基因空间表达模式分析
• 检测青蒿素途径中几个重要化合物含量,获得不同株系中差异代谢物积累情况
• 着重ABP途径中各个基因家族的变异、拷贝和表达情况,查看是否能与代谢物积累相关联
• 获得青蒿素代谢关联基因,对其表达谱、基因特征进行详细分析,阐述青蒿素合成、调控以及差异积累差异的原因。
• 获得选种标记
6.阅读中问题
研究背景
• 使用青蒿素复方疗法,许多国家在消灭疟疾方面取得了进展。青蒿的青蒿素含量变化很大,通常在 0.01% 到 1% 之间。在农业青蒿素供应不稳定的情况下,培育青蒿素含量高的优良种质是缓解青蒿素药物需求压力的关键环节。 因此,青蒿基因研究的重要性怎么强调都不为过。
• 由于青蒿的高杂合性和重复性,目前只有遗传图谱 (Graham et al., 2010) 和支架 N50 长度为 104.86 kb (Shen et al., 2018) 的基因组草图可用。 缺乏完整准确的基因组结构,阻碍了优良品种的分子辅助育种和青蒿素代谢工程。
• 青蒿素主要合成并储存在叶表面的腺体分泌毛状体(GSTs)中; 然而,非 GST 细胞也能够产生青蒿素。
• 排除内过氧化物桥的最后脱氢,青蒿素生物合成途径(ABP)已得到很好的阐明。青蒿素由异戊烯基焦磷酸经法尼基焦磷酸( FPP )合成,在紫穗槐-4,11-二烯合酶( ADS )作用下转化为紫穗槐-4,11-二烯,这被认为青蒿素合成第一步。通过细胞色素 P450 单加氧酶 (CYP71AV1) 的催化将 amorpha-4,11-二烯三步氧化为青蒿酸、青蒿醇和青蒿醛。青蒿醛也通过双键还原酶2 ( DBR2 )交替还原为双氢青蒿素醛。该途径的后期由乙醛脱氢酶1 ( ALDH1 ) ( Teoh等, 2009)介导,催化青蒿醛和双氢青蒿素醛氧化,分别生成青蒿酸和二氢青蒿酸(附图1)。青蒿素生产的最后阶段被认为是光诱导的非酶促光化学氧化过程反应。
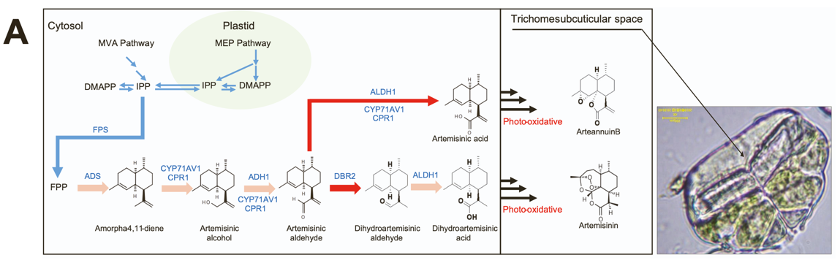 附图1青蒿素合成
附图1青蒿素合成
•
• 叶面积、腺毛密度和植株鲜重是提高青蒿素产量的目标性状。此外,高温胁迫、UV - B辐射、UV - C辐射等外源因子以及JA、ABA、SA等植物激素的处理均能显著提高青蒿素的合成。
• 多个转录因子( transcription factors,TFs )可以上调青蒿素的合成,包括AaWRKY1 、AaORA、AabHLH1、AabZIP1、AaMIXTA1、AaMYB2和AaMYC2-like。然而,青蒿素生物合成的遗传学方面的许多重要问题仍未得到解答。这些包括(a)ABP基因的结构、数目、基因组位置和进化历史;(b) ABP基因的遗传特性与青蒿素生产之间的关系;(c)基因组中ABP基因顺式作用元件/反式作用因子的全息图。
• 为了回答这些问题,本研究利用PacBio Hi-Fi reads、Bionano光学图谱和Hi-C数据从HAN1 (高青蒿素菌株,干重的1%)和LQ-9 (低青蒿素品系,干重0.11 %) 中获得了近乎完整的染色体水平的阶段基因组。此外,对36个品种进行了重测序。黄花蒿具有丰富的遗传多样性,ADS拷贝数与青蒿素含量呈正相关。这种高质量的黄花蒿基因组信息为青蒿素的生物合成及其调控提供了新的见解,加深了我们对黄花蒿遗传背景的理解,为黄花蒿分子育种奠定了基础。
结果分析
基因组组装和单倍型染色体构建
• LQ-9和HAN1的青蒿素含量分别为干重的0.11±0.01%和1.05±0.09%,与页面GSTs的密度密切相关。
• 104X Illumina short PE150 reads, 35X PacBio Hi-Fi reads, 520X Bionano optical map data, and 187X Hi-C data were generated for LQ-9, and 85X Illumina short PE150 reads, 25X Hi-Fi reads, and 255X Bionano data were generated for HAN1。
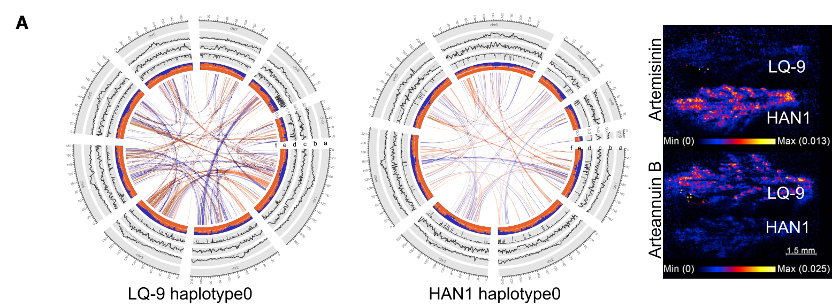
• 四个单倍型基因组平均大小为1.5Gb,平均预测杂合度2.1%。更长的scaffold为haplotype0,短且碎的为haplotype1。

• 当使用单个单倍型作为参考时,观察到比使用两个单倍型作为参考时显着更高的 SNP 密度
> 作者解答:SNP这个问题我们当时没有参考其他工作,它基于一个推理,就是如果只用一套倍型做参考,那么另一套的reads会报告SNP。如果用两套倍型的基因组,在一个区段未发现连续的SNP,说明拼接杂合低,基因组准确
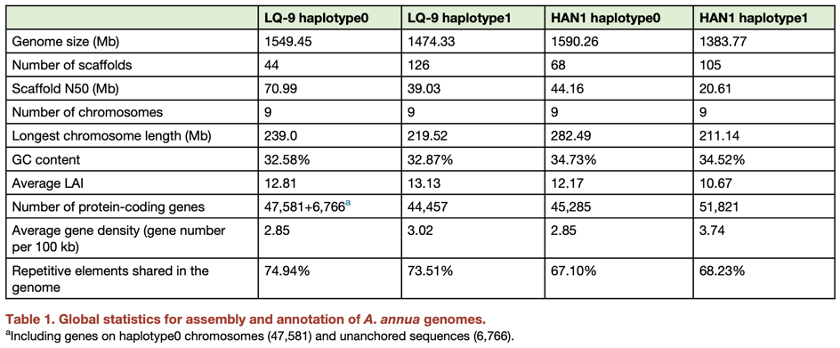
表1:基因组统计
2. Evolution of the A. annua genome
黄花蒿基因组进化
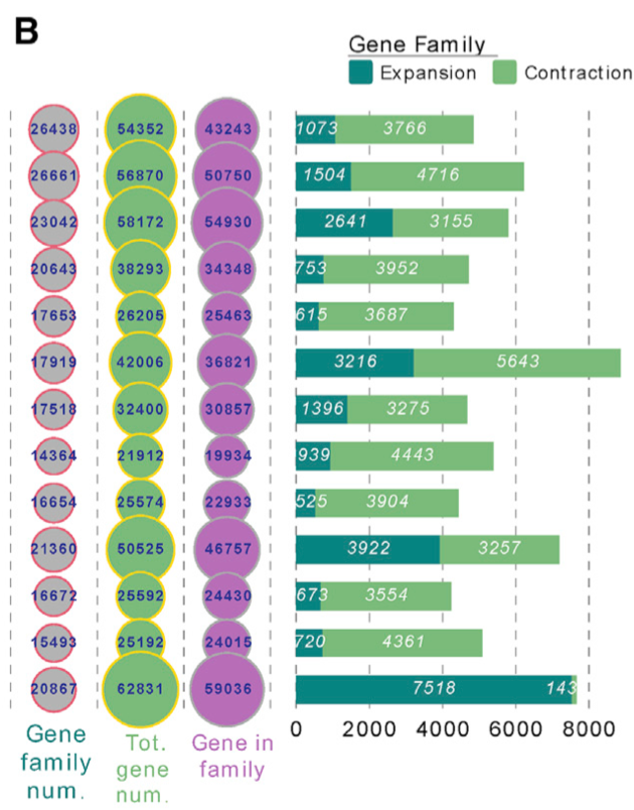
• 通过比较黄花蒿LQ-9单倍型0基因组与12种代表性植物基因组,分析了基因家族的进化动态。在黄花蒿基因组中共鉴定出26,438个候选基因家族,其中3,125个为黄花蒿特有,3,776个为所有13个物种共有。
• 在黄花蒿中,共有1,073个基因家族似乎已经扩张(图2B ),包括R -芳樟醇合成酶活性( GO : 0034008 ,富集p = 1.5e ~ ( 14 ))和香叶基二磷酸代谢过程( GO : 0033383 ,富集p = 5.78eα14)
• 利用21个共有的单拷贝基因构建了一个系统发育树。黄花蒿和菊花脑的分歧时间为11.43 Ma,而黄花蒿和向日葵的分歧时间为37.96 Ma(图2A)
• 通过基因组内比较,在黄花蒿LQ9单倍型0基因组中共发现418个共线性片段,包括2 596个旁系同源基因对,占基因总数的9.55%。
• 共线性旁系同源基因(图2C)之间的同义替换率( Ks )与共线性区段中存在的33个区域之间的正常密度分布,以及咖啡、生菜和向日葵基因组之间的共线性区段(图2D ),表明除了全基因组三倍化( WGT ) g (在咖啡基因组中显示)外,在菊科物种分化之前的58.12 Ma (平均Ks 1.07),黄花蒿进化过程中发生了一次WGT事件。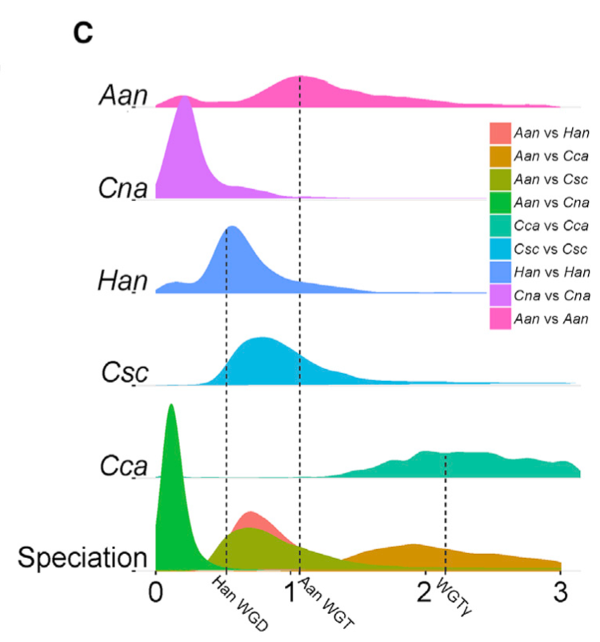
•
• 进一步分析发现,WGT后基因保留(涉及403个基因)和串联重复(涉及基因723个)占扩增到的基因家族(共计1,058个基因)的46.4%,说明这些现象在基因家族扩张中起主要作用。
3.Abundance of genetic diversity in A. annua
黄花蒿的遗传多样性丰富
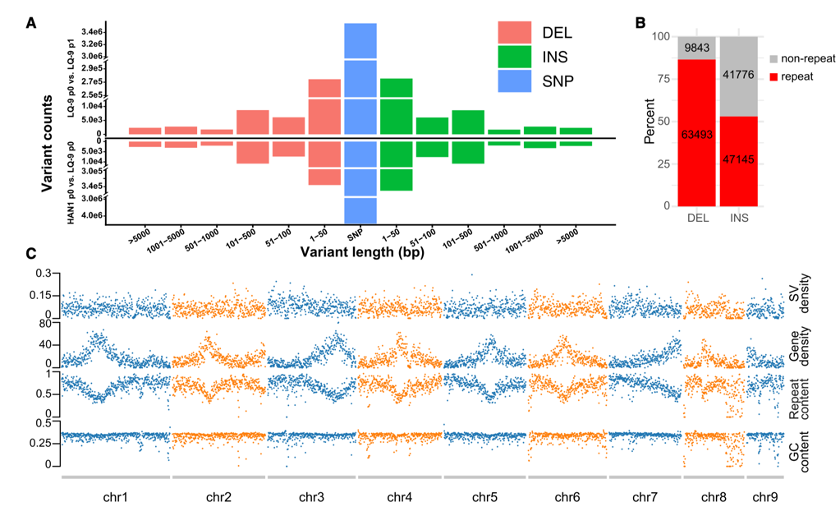
• 为了识别序列变异,使用Minimap2将LQ-9haplotype1、HAN1haplotype0、HAN1 haplotype1和HuHao的最终组装映射到LQ-9 haplotype0组装,检测SNP,和结构变异(INDEL和SVs)。
• SNP的平均密度为0.73个/ 100 bp。大多数SV(结构变异)的长度在2 kb以内(图3A ),约68%的SV与重复序列重叠(图3B )。然而,SV倾向于均匀分布在基因组上,而与基因密度或重复密度没有显著关系(图3C )。
•
• 值得注意的是,HAP和LAP株系保持着各种单萜和倍半萜的特定积累模式。编码单萜和倍半萜生物合成关键酶的TPS基因家族在单倍型或株系间也表现出基因数量、结构和表达谱的多样性。
4.Spatial expression profiles of A. annua genes
黄花蒿基因的空间表达谱
• 植物化学(Phytochemical )研究主要集中在黄花蒿叶片,因为青蒿素及其相关化合物主要在叶片的GST和非GST细胞中合成。成像质谱(Imaging mass spectrometry)已被用于检测次级代谢产物的原位积累模式。
• 解吸电喷雾电离质谱( DESIMS )成像显示的信号响应表明,HNA1叶片中青蒿素(artemisinin)的原位积累高于LQ-9(下图)。相比之下,青蒿乙素(arteannuin B)在HAN1叶片中的原位积累量低于LQ-9叶片。UPLC-ESI-QQQ-MS也获得同样的结果。

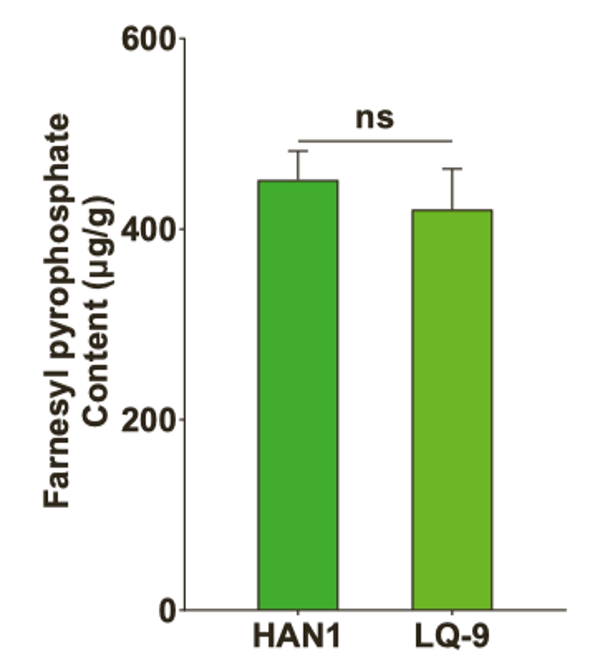
• GSTs中的FPP含量在两株系(附图1C)之间没有显著差异,表明不同的机制负责ABP基因的表达和转录调控。
• 基于HAN1和LQ-9 (附表1)的4个组织(根、茎、叶和花)和31个公开数据集(所有样品来自根、茎、叶或花的组织,在Illumina HiSeq平台上测序;详见附表16)的24个RNA测序数据,对ABP的表达和转录调控提供了新的见解。
• 总共有14,827个基因被发现在来自两个菌株的四个组织中差异表达(log2[fold change]>=2,调整的p = 1e-6),其中6,820个基因表现出高度组织特异性表达( TAU index>0.9)。
• 大部分ABP基因在花或叶中高表达,而在CYP71AV1基因和DBR2不同拷贝中的表达模式不同。CYP71AV1基因的一个拷贝在叶和花中高表达,另一个拷贝在根和茎中高表达。 DBR2.1在所有组织中均有表达,DBR2.2在叶片和花中高表达(图4A)。
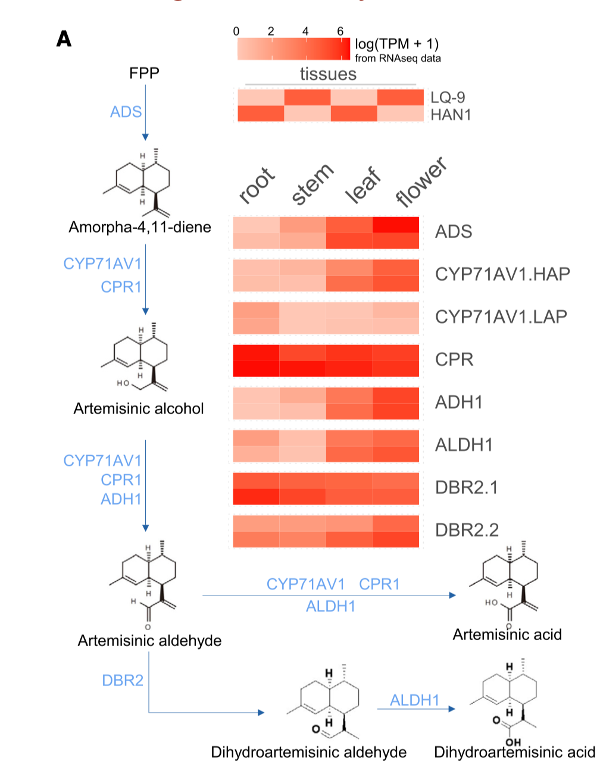
• 在叶片中,LQ-9和HAN1之间检测到3,542个DEGs。在这些差异表达基因中,ADS和ADH1的所有拷贝、DBR2的1个拷贝和CYP71AV1基因的1个拷贝均差异表达(附图15),表明这些位点可能在青蒿素积累中起关键作用。值得注意的是,所有的ABP基因都至少有一个拷贝,在花中高表达,这与以前的研究报告的花中GSTs的高密度相一致。
花中也产生青蒿素?
• WGCNA分析显示,所有已知ABP基因的至少一个拷贝聚在同一个模块(pink module),其中包含1,744个基因并且和青蒿素含量成正相关。粉色模块中的基因表现出与蜡质合成相关的显著功能富集,被认为与GST形成和青蒿素生物合成相关。
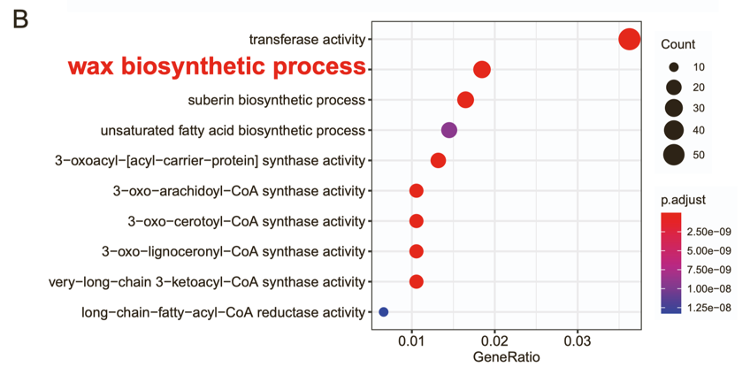
• 粉红色模块包括来自14个家族(如MYB和锌指TFs等)的63个转录因子,这些转录因子与青蒿素生物合成途径中的基因共表达。其中2个转录因子与ORA具有较高的蛋白序列一致性,2个转录因子GSW已被证实调控青蒿素生物合成。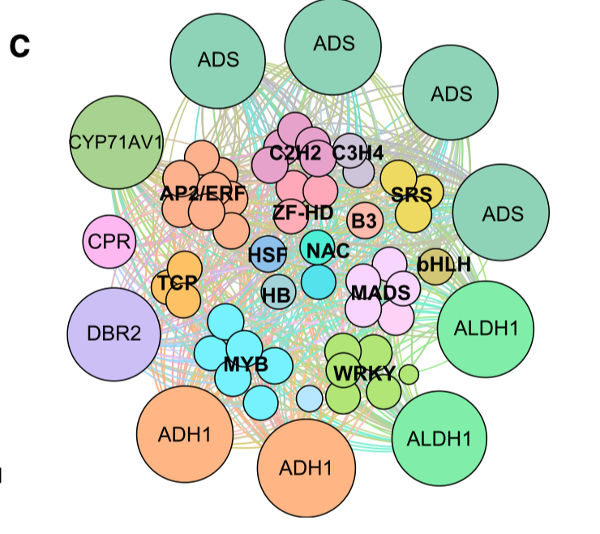

5.Structural characteristics and expression profiles of ABP genes
ABP基因的结构特征和表达谱
• ABP基因具有丰富的多样性,基因的表达量变化较大(图4A)。基因复制是ABP基因中常见的进化事件,因为在四个单倍型图谱(附图17)中,所有ABP基因都至少检测到一个相邻或分散的拷贝。这些基因的拷贝数从2到6不等,串联基因簇的长度从9.45 kb到121.49 kb不等(表2)。
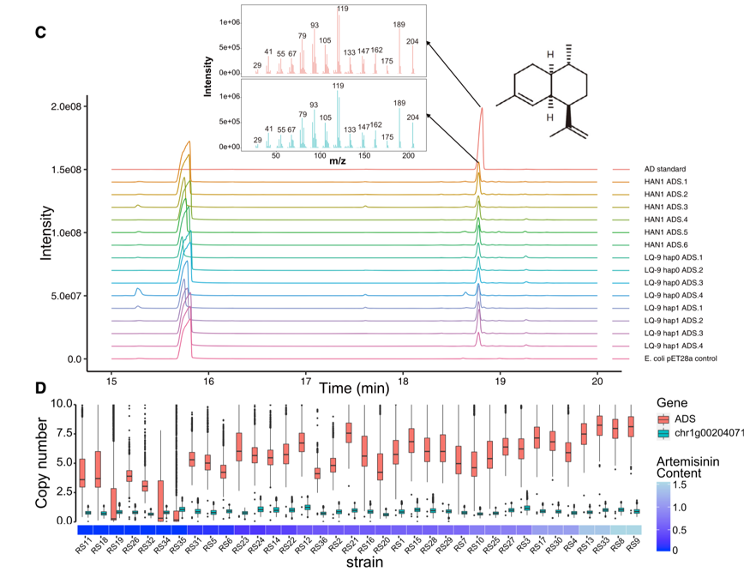
• 在每个基因的拷贝之间观察到高的全局序列一致性( 85.66% ~ 100%),表明它们重复后的分歧时间很短。ABP中的第一个关键基因ADS显示出不同拷贝数的完整基因结构,从LQ-9单体型0中的4个到HAN1单体型1中的6个,这与使用Bionano分子(图5A和5B)的检测结果一致。
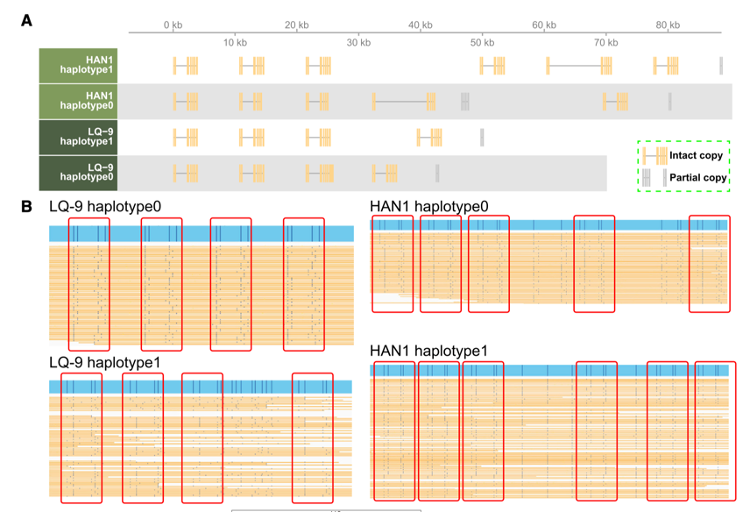
是串联重复
• ABP基因在HAN1和LQ-9 (附图15)叶片中存在差异表达。具体来说,结合RNA-seq和IsoSeq数据中所有拷贝的表达,ADS在HAN1中的表达高于LQ-9叶片。考虑到LQ-9和HAN1中GSTs的FPP含量没有显著差异,ADS的差异表达可能导致紫穗槐- 4,11 -二烯含量(附图1)的差异,从而影响两株系从FPP到整体青蒿素生物合成的代谢通量。
• 为了鉴定ABP基因拷贝之间的表达谱,对LQ-9的根、叶和花组织以及HAN1的叶组织进行了4组Iso-Seq数据(采用PacBio HiFi测序)。基于从Iso-Seq数据获得的全长转录本,在所有ADS拷贝的CDS中总共鉴定出23个SNP。所有的ADS拷贝被SNP位点分为14种类型。
• 在Iso - Seq数据集(图4B )中发现所有单独的ADS拷贝都被转录,并且具有催化FPP生成AD的酶活性(图5C),表明扩增的ADS拷贝增强了其转录。HAN1中ADS较高的转录水平可能是由于其较高的基因拷贝数。对不同青蒿素含量的重测序样品进行了相对深度分析(图5D)。在36个(青蒿素含量= ADS拷贝数3 0.13 + 0.02 ; Pearson相关性0.75 ; R2 = 0.57 ; p = 1.08eα7)个体中观察到青蒿素含量与ADS拷贝数之间存在很强的线性正相关。ADS的拷贝数可能是区分高、低青蒿素化学型的关键特征,这将使ADS成为黄花蒿遗传育种的可靠标记。
• ABP中的另一个关键基因DBR2在每个单倍型中有两个拷贝( DBR2.1和DBR2.2)。CDS分别在DBR2.1和DBR2.2单倍型之间保守。这两个拷贝的蛋白序列存在差异,在C端(附图19)缺失了7个氨基酸的延伸。DBR2 (DBR2.2)的删除版本的功能已被报道。DBR2的两个拷贝的表达谱是不同的(图4B和补充图14)。在IsoSeq数据集中,DBR2.2的最高转录本水平在HAN1的叶片中,在LQ-9叶片数据集中没有检测到DBR2.2转录本(图4B )。
讨论
名词解释
• ABP:artemisinin biosynthetic pathway,青蒿素生物合成途径
• GSTs:glandular secreting trichomes腺毛状体,青蒿素主要合成并储存在叶表面的腺体分泌毛状体(GSTs)中; 然而,非 GST 细胞也能够产生青蒿素
• Imaging mass spectrometry :成像质谱
• Phytochemical:植物化学
• FPP:farnesyl pyrophosphate,法尼基焦磷酸。青蒿素由异戊烯基焦磷酸经法尼基焦磷酸( FPP )合成,在紫穗槐-4,11 -二烯合酶( ADS )作用下转化为紫穗槐- 4,11 -二烯。
• artemisinin:青蒿素
• TAU index
文献参考
[1] LIAO B, SHEN X, XIANG L, et al. Allele-aware chromosome-level genome assembly of Artemisia annua reveals the correlation between ADS expansion and artemisinin yield [J]. Mol Plant, 2022, 15(8): 1310-28.
请大家多多评论,如果有理解不正确的地方希望指正
https://blog.sciencenet.cn/blog-3422766-1359150.html
上一篇:PolyGembler:多倍体和高杂合植物基因组遗传作图新方法