ВЉЮФ
ДњаЛбЇШЫ--Nature MetabolismЃКИпжЌвћЪГКѓЃЌЮвЕФСмАЭЙмТЉСЫ...
||
ДњаЛбЇШЫ
Nature MetabolismЃКИпжЌвћЪГКѓЃЌЮвЕФСмАЭЙмТЉСЫ...

зЋЮФ | СѕЫЌ ВмгёЯу ИпвнФў Сѕаугё РюЙњЧП
БрМ | УЯУРбў
аЃЖд | РюЙњЧП
БГОАНщЩм
ЫцзХЩњЛюЫЎЦНЕФЬсИпЃЌЗЪХжЯжвбГЩЮЊгАЯьШЋЧђЕФТ§адДњаЛадМВВЁЁЃвХДЋвђЫигыЛЗОГвђЫиЙВЭЌзїгУгеЗЂЗЪХжЁЃвЛЗНУцЃЌЪнЫи(leptin)ЁЂНтХМСЊЕААзЃЈUCP1ЃЉЁЂЩёОыФYЃЈNPYЃЉЁЂжзСіЛЕЫРвђзг-ІСЃЈTNF-ІСЃЉЕШЗЪХжЯрЙиЛљвђЕФЭЛБфЛсв§ЗЂвХДЋадЗЪХжЃЛСэвЛЗНУцЃЌИпжЌЩХЪГЁЂдЫЖЏШБЗІЕШФПЧАГЃМћЩњЛюЗНЪНвВМгОчЗЪХжЕФЗЂЩњЁЃбаОПЗЂЯжЃЌЫцзХЗЪХжЕФЗЂЩњЗЂеЙЃЌЛњЬхвШЕКЫиЗжУкМѕЩйЃЌИЮдрЁЂжЌЗОЕШзщжЏЖдЦЯЬбЬЧКЭвШЕКЫиЕФУєИаадЯТНЕЃЈМДВњЩњвШЕКЫиЕжПЙЃЉЃЌНјЖјЗЂеЙЮЊ2аЭЬЧФђВЁЕШДњаЛадМВВЁЁЃ
ШЫЬхСмАЭЯЕЭГЖдгкЬхвКЦНКтКЭУтвпЗРЛЄЦ№зХжСЙиживЊЕФзїгУЃЛСмАЭЯЕЭГНсЙЙКЭЙІФмЕФЮЩТвГЃЪЧЕМжТЛђМгжиАЉжЂЁЂздЩэУтвпКЭбзжЂадМВВЁЃЌаФФдбЊЙмМВВЁвдМАЩёОЭЫааадМВВЁживЊвђЫижЎвЛЁЃСмАЭЯЕЭГгыжЌЗОДњаЛвВгаНєУмСЊЯЕЃЌР§ШчдкСмАЭЫЎжзЃЈгЩСмАЭЙмЛиСїЙІФмЪмзшЕМжТЃЉЛМепКЭОпгавьГЃСмАЭЯЕЭГЕФзЊЛљвђаЁЪѓжаЃЌжЌЗОзщжЏЛсдкСмАЭЙмЕФБЁШѕВПЮЛЃЈСмАЭЩјТЉВПЮЛЃЉОлМЏЃЌЯдЪОЗЪХжгыСмАЭЯЕЭГЙІФмЪмЫ№гаЙиЁЃБШШчЃЌдквХДЋадЬЧФђВЁЃЈdb/dbЃЉаЁЪѓКЭЮЙЪГИпЙћЬЧвћЪГЕФЗЪХжДѓЪѓЬхФкЃЌГІЯЕФЄжЌЗОзщжЏЙ§ЖШЖбЛ§ЃЌГІЯЕФЄСмАЭЙмЙІФмеЯАЃЌСмАЭЙмЭЈЭИадКЭЪеЫѕадЗЂЩњСЫИФБфЁЃГІЯЕФЄжЌЗОзщжЏЕФЙ§ЖШЖбЛ§ЛсЕМжТЦфжЌЗОЫсКЭжЌЗОвђзгЙ§ЖШЪЭЗХЃЌНјЖјДйНјЛњЬхВњЩњвШЕКЫиЕжПЙЁЃдкШЫЬхИЙЧЛФкЃЌГІЯЕФЄжЌЗОзщжЏАќЙќзХГІЯЕФЄСмАЭЙмКЭСмАЭНсЃЌСмАЭЭЈЙ§етаЉСмАЭЙмКЭСмАЭНсЭљРДгкГІКЭГІЯЕФЄЁЃДЫЧАKuanЕШШЫбаОПЗЂЯжЃЌГІЯЕФЄКЭжмЮЇСмАЭЙмЕФЙЬгаЭЈЭИадЪЙСмАЭГЩЗжФмЙЛЗжВМЕНГІЯЕФЄжЌЗОзщжЏЁЃФЧУДСмАЭЙІФмеЯАгыГІЯЕФЄжЌЗОзщжЏЕФЖбЛ§ЃЌЗЪХжЁЂвШЕКЫиЕжПЙгаЪВУДЙиЯЕФиЃП
аЁБрзЂЃК1ЁЂСмАЭЙмЙІФмеЯАКЭжЌЗОзщжЏОлМЏЃК
ЃЈ1ЃЉГЃМћЕФЪЧЪжБлКЭЭШВПСмАЭвКХХГіЪмЫ№Жјв§Ц№ЕФЫЎжзЃЌЖдгкЭэЦкСмАЭЫЎжзЮЊСЫМѕЧсЪжБлКЭЭШВПжзеЭвЛАуНјааГщжЌжЮСЦЃЌЯТЭМзѓВрЪЧгвжЋГщжЌЃЌжаМфЪЧГщжЌ6ИідТЃЌгвЭМГщжЌ12ИідТЁЃ

ЃЈ2ЃЉгаЮФЯзбаОПБэУїApelinЭЈЙ§діЧПСмАЭКЭбЊЙмЭъећадвжжЦвћЪГгеЕМЕФЗЪХжЃЌЖјApelinЧУГ§аЁЪѓдкHFDЬѕМўЯТЦфИЙЙЩЙЕжЌЗОжЌжЪЖбЛ§ЃЌжЌЕЮБфДѓШчЯТЭМЫљЪОЁЃ

2ЁЂСмАЭЙмЭЈЭИадКЭЩьЫѕадШчКЮЙлВьЃП
ЃЈ1ЃЉdb/dbаЁЪѓСмАЭЙмЭЈЭИадИФБфЕФЪЕбщЃКЪзЯШЃЌНЋЯдЮЂОЕЯТЖЈЮЛЪеМЏЕФГІЯЕФЄСмАЭЙмзЊвЦЕНвЛИіИєРыЕФШнЦїжаЃЌВЂНЋЦфСЌНгЕНВЃСЇЮЂСПвЦвКЙмЩЯЃЌДЫвЦвКЙмСНВрКЌгаЯрЭЌЕФШмвКЃЌЕЋЦфжавЛВрКЌгаТЬЩЋгЋЙтБъМЧЕФАзЕААзBSAЁЃЫцзХЪБМфЕФЭЦвЦЃЌгЋЙтBSAДЉЙ§СмАЭЙмБкНјШыШмвКЃЌЕМжТЙтЖШМЦЕчбЙж№НЅГЪЫЋЯпаддіМгЃЌНЋЙрСївКИФЮЊЮДБъМЧЕФBSAЛсЧхГ§ЫљгагЋЙтЃЌЙтЖШМЦЕчбЙЛжИДжСЛљЯпЃЌИљОнЗЦПЫЕквЛЖЈТЩМЦЫуBSAЕФЭЈСПЃЌвдДЫМьВтСмАЭЙмЕФЭЈЭИадДѓаЁЁЃ
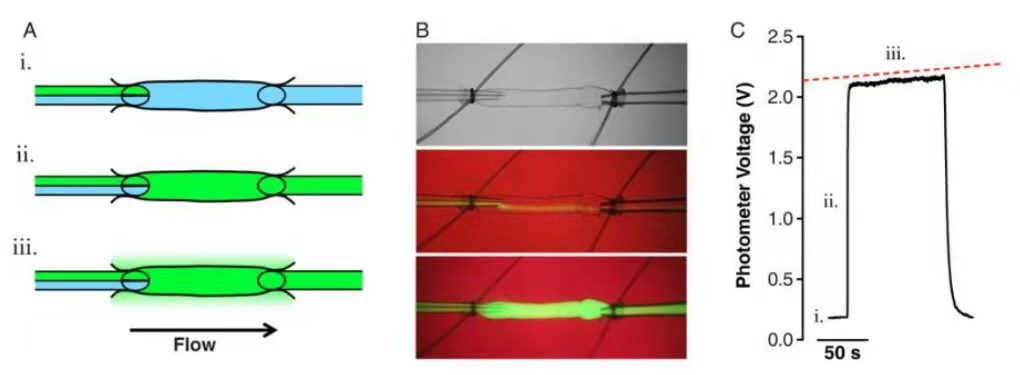
ЃЈ2ЃЉСмАЭЙмЪеЫѕадИФБфЕФЪЕбщЃКЪзЯШЃЌНћЪГ16аЁЪБЃЌШЛКѓТщзэЖЏЮяЃЌЦЪПЊИЙЧЛЃЌЭтжУ6~7cmГЄЕФГІЯЕФЄДВЛЗЁЃНЋГІЯЕФЄДВеЙЦНЃЌДгжмЮЇзщжЏжазаЯИЗжРыГі2ЬѕГІЯЕФЄСмАЭЙм(MLV)ЃЌЧхГ§ЫљгажЌЗОзщжЏШЛКѓгУгкСмАЭЙмЪеЫѕадЪЕбщЁЃСмАЭЙмЪеЫѕадЪЕбщШнЦїСЌНгЕНЕЅЖРЕФАзЕААзВЙГфЩњРэбЮЫЎШмвКбЙСІШнЦїжаЃЌШЛКѓНЋУПИіСмАЭЙмЗжБ№АВзАдкГфТњAPSSЛКГхвКЕФвЧЦїжаЃЌЕїНкЕНЦфДѓдМЕФдЮЛГЄЖШЃЌВЂдЪаэдк1cmH2OЕФПчБкбЙСІЯТдкAPSSжаБЃГжЦНКтЃЌжБЕНЙлВьЕНЯрадЪеЫѕЁЃбЊЙмБЉТЖдк1ЁЂ3КЭ5cmH2OЕФПчБкбЙСІЯТЃЌМЧТМЪеЫѕЛюЖЏИї5ЗжжгЁЃзюКѓЃЌЭЈЙ§НЋдЁВлЧаЛЛЕНЮоИЦAPSSЃЌМЧТМУПИіПчБкбЙСІЯТЕФБЛЖЏСмАЭЙмжБОЖЁЃ
зюНќЕФNature MetabolismЩЯЯпСЫEnyuan CaoЕШШЫЙигкHFDвћЪГгеЕМГІЯЕФЄСмАЭЙмЙІФмеЯАЃЈжївЊЪЧГІЯЕФЄСмАЭТЉШыГІЯЕФЄжЌЗОзщжЏЃЉЃЌНјЖјДйЪЙвШЕКЫиЕжПЙЕФЯрЙибаОПЃКЁАMesenteric lymphatic dysfunction promotesinsulin resistance and represents a potential treatment target in obesityЁБЁЃБОЦЊбаОПБэУїЃЌСмАЭжаСмАЭЯИАћЕФCOX-2КЭVEGF-C-VEGFR3аХКХЭЈТЗПЩвдЭЈЙ§ДйНјЗЪХжЯрЙиЕФГІЯЕФЄСмАЭЙІФмеЯАЃЌЕМжТЛњЬхВњЩњвШЕКЫиЕжПЙЃЛЪЙгУШћРДЮєВМЧАвЉЃЈCele-ProЃЉАаЯђвжжЦСмАЭжаЕФCOX-2БэДяЃЌПЩгааЇФцзЊГІЯЕФЄСмАЭЙІФмеЯАЃЌИФЩЦЗЪХжВЂПижЦЛњЬхбЊЬЧЁЃИУбаОПНсЙћНвЪОСЫеыЖдЗЪХжЯрЙиГІЯЕФЄСмАЭЙІФмеЯАЪЧжЮСЦДњаЛадМВВЁЕФвЛжжЧБдкжЮСЦбЁдёЁЃЃЈаЁБрзЂЃКдкБОЦЊЮФеТжаЃЌФкдржЌЗОзщжЏ(VAT)ЬижИАќЙќзХГІЯЕФЄСмАЭЙмКЭСмАЭНсЕФжЌЗОзщжЏЃЌСмАЭЭЈЙ§ЫќУЧдкГІКЭГІЯЕФЄдЫЪфЁЃЃЉ

1.HFDвћЪГгеЕМГІЯЕФЄСмАЭЙмЙІФмеЯАЁЃ
2.COX-2КЭVEGF-CКЌСПЩ§ИпЃЌЕМжТвьГЃСмАЭЙмдіЩњКЭСмАЭвКЩјТЉЁЃ
3.вжжЦVEGF-CПЩгааЇЛжИДСмАЭЙмЙІФмЁЃ
4.ШћРДЮєВМЧАвЉЃЈCele-ProЃЉгааЇФцзЊHFDвћЪГгеЕМГІЯЕФЄСмАЭЙмЙІФмеЯАЁЂИФЩЦаЁЪѓЗЪХжВЂПижЦбЊЬЧЁЃ
баОПНсЙћ
1. HFDгеЗЂЗЪХжЪБЃЌГІЯЕФЄСмАЭЙмЗЂЩњжиЙЙ
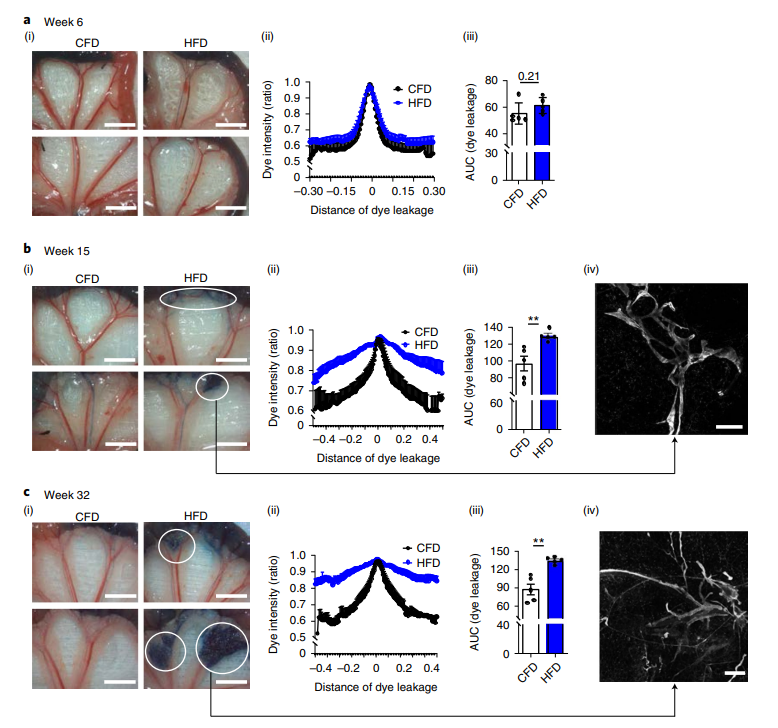
баОПШЫдБЪзЯШЖдаЁЪѓЮЙЪГHFDЛђЖдеевћЪГЃЈCFDЃЌМДе§ГЃвћЪГЃЉ6ЁЂ15ЁЂ23КЭ32жмЃЌВЂЖдетаЉаЁЪѓЕФСмАЭЙмЁЂГІШоУЋКЭжЌЗОЯрЙиСмАЭДиЃЈFALCsЃЉЕФНсЙЙНјааСЫБШНЯЁЃНсЙћЯдЪОаЁЪѓHFDЮЙбј15жмКѓЃЌЪЎЖўжИГІжаЕФШоУЋКЭШщУгЙмУїЯдБфПэЃЌШоУЋБфЖЬЃЌШщУгЙмГЄЖШЛљБОВЛБфЃЈРЉеЙЪ§ОнЭМ1a-eЃЉЃЛЫцзХHFDЮЙбјЪБМфЕФдіМгЃЌVATжаГІЯЕФЄСмАЭЙмж№НЅГіЯжЗжжЇЁЃдкHFDЮЙбјЕФЕк6жмЃЌСмАЭЙмЗжжЇЕФдіМгВЛЯджјЃЌдкHFDЮЙбјЕФЕк15жмЃЌСмАЭЙмЗжжЇЯджјдіМгЃЌдкHFDЮЙбјЕФЕк32жмЃЌСмАЭЙмЗжжЇдіМгЕиИќЮЊЯджјЃЌЧвСмАЭЙмаЮЬЌНЯЮЊЮЩТвЭфЧњЃЌВЂЗЧбизХСмАЭСїЗНЯђЩњГЄЃЈЭМ1aЃЌbЃЉЁЃЭЌЪБЃЌбаОПШЫдБвВЖде§ГЃШЫКЭЗЪХжЛМепVATжаЕФСмАЭЙмНјааСЫМьВтЃЌЗЂЯжЗЪХжЛМепЕФСмАЭЙмЗжжЇдіМгЃЌVATжажБСмАЭЙмМѕЩйдМ35%ЃЌгиЧњСмАЭЙмдіМгдМ50%ЃЌСмАЭЙмаЮзДЮЩТвЃЈЭМ1cЁЂdКЭРЉеЙЪ§ОнЭМ1hЃЉЃЌЫЕУїдкЗЪХжЕФЗЂЩњЙ§ГЬжаЃЌСмАЭЙмЕФаЮзДКЭНсЙЙЗЂЩњжиЫмЁЃ
жЎЧАВЂЮДгаЙ§ЗЪХжЧщПіЯТFALCsЕФбаОПБЈЕРЁЃдкБОбаОПжаЃЌбаОПШЫдБЗЂЯжаЁЪѓОЙ§HFDЮЙбј15жмКЭ32жмКѓЃЌFALCsЕФЪ§СПЯджјдіМгЁЃЯрСкЕФFALCЃЌFLACгыVATЕФЗжжЇСмАЭЙмЯрЛЅСЌНгЃЌаЮГЩИДдгЕФСмАЭЭјТчЃЈЭМ1e-gЃЌРЉеЙЪ§ОнЭМ1iЈCmЃЉЃЌНјвЛВНЫЕУїHFDгеЕМЗЪХжЪБЃЌVATжаСмАЭЙмЛсЗЂЩњжиЙЙЁЃ
ЭиеЙдФЖС
СмАЭЙмбаОПЗНЗЈбЇ
БОЮФРћгУСЫЖржжЗНЗЈРДЖдСмАЭЙмНјааСЫбаОПЁЃ
УтвпгЋЙтШОЩЋБъМЧLYVE1(СмАЭФкЦЄЯИАћЭИУїжЪЫсЪмЬх1)РДЙлВтСмАЭЙмЕФаЮЬЌЃЛРћгУYFPгЋЙтФЩУзЧђИЙЧЛзЂЩфЙлВьFLACsЕФаЮЬЌЁЃ
вСЮФЫМРЖШОСЯЛюЬхзЂЩфЖдСмАЭЩјТЉЧщПіНјаабаОПЁЃОпЬхЗНЗЈЮЊЃКИЙВПЧаПкЃЌНЋаЁГІЗГіЬхЭтВЂгУЮТPBSШѓЪЊЁЃЗжРыГі4-5РхУзЕФЪЎЖўжИГІЛђПеГІЖЮЃЌЦфжаАќКЌPeyer'sСмАЭНсЁЃНЋ5ul 10%вСЮФЫМРЖШОСЯзЂШыPeyer'sСмАЭНсЕФЩЯВуЁЃдкзЂЩфКѓЕк2,5,10,20КЭ30ЗжжгМЧТМвСЮФЫМРЖШОСЯЭЈЙ§ГІЯЕФЄСмАЭЙмв§СїВЂЩјШыжмЮЇVГІЯЕФЄжЌЗОзщжЏЁЃЪЙгУЯрЛњВЖЛёЭМЯёЁЃДгСмАЭЙмжааФЯђЭтВтСПШОСЯЧПЖШжСжЌЗОзщжЏЁЃЬсШЁВЂзЊЛЛШОСЯЧПЖШЪ§ОнвдеыЖдзюДѓаХКХЧПЖШНјааБъзМЛЏЁЃ
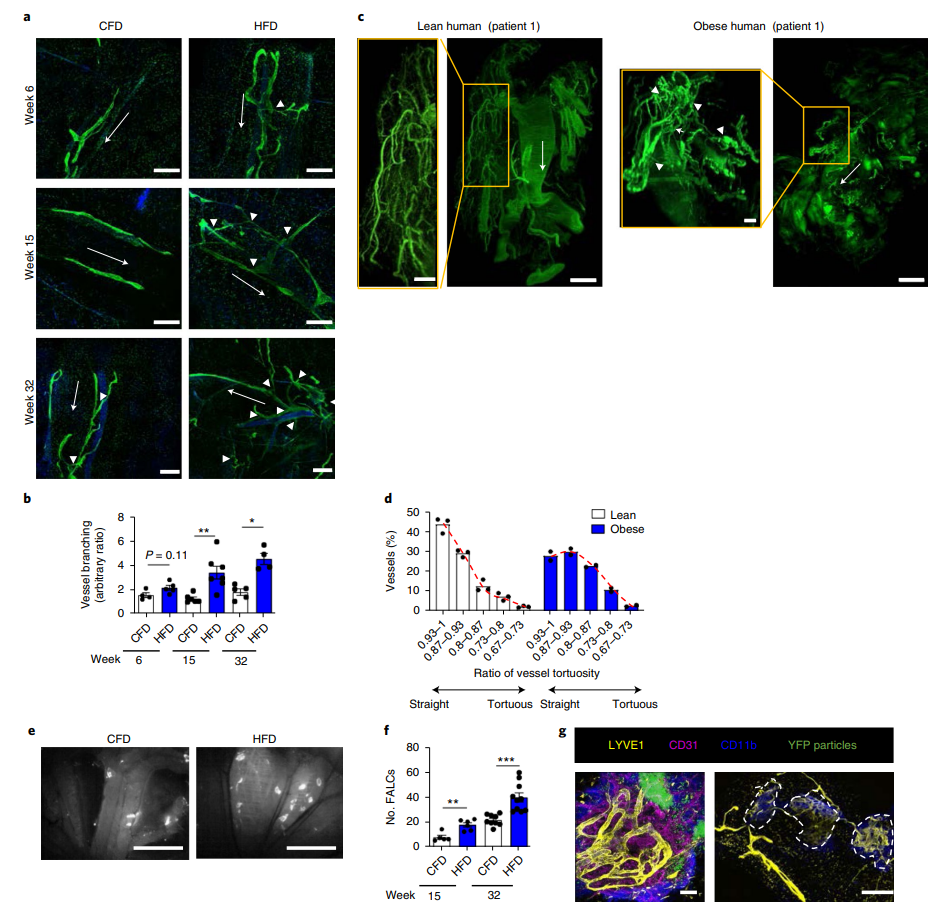
ЭМ1. HFDгеЗЂЕФЗЪХжгыНјааадГІЯЕФЄСмАЭЙмжиЫмгаЙи
РЉеЙЪ§ОнЭМ1.дкГІЯЕФЄжЌЗОзщжЏжаЃЌHFDЯрЙиЕФГІШоУЋКЭШщУгЙмЁЂжЌЗОЯИАћКЭFALCЕФБфЛЏ
2. HFDгеЗЂЗЪХжЪБЃЌГІЯЕФЄСмАЭТЉжСГІЯЕФЄжЌЗОзщжЏ
ЮЊСЫЦРЙРГІЯЕФЄСмАЭЙмЕФв§СїКЭзЊдЫЙІФмЃЌбаОПШЫдБНгЯТРДЖдЮЙЪГCFDКЭHFDЕФаЁЪѓНјааСЫвСЮФЫМРЖШОСЯСмАЭЙмдьгАЪЕбщЃЈаЁБрзЂЃКвСЮФЫМРЖШОСЯСмЕФзїгУЪЧЮЊСЫПЩЪгЛЏГІЯЕФЄСмАЭЙмЩјТЉЧщПіЁЃЫќЪЧвЛжжПЩвдЛюЬхзЂЩфЕФШОСЯЃЌвСЮФЫМРЖгыбЊвКжаЕФбЊНЌЕААзгаКмИпЕФЧзКЭСІЃЌвЛАудкЩёОбЇПЦбаОПжаБЛгУзїЪОзйМСЙлВьбЊФдЦСеЯЕФЭъећадЁЃДЫЭтЃЌвСЮФЫМРЖЛЙФмЙЛдкЬхЭтЯдЮЂОЕЯТЧјЗжЫРЯИАћгыЛюЯИАћЃЌДЫЪБзїгУРрЫЦгкЬЈХЮРЖЃЉЁЃбаОПШЫдБНЋШОСЯзЂЩфЕНГІеГФЄжаЃЌЗЂЯжЫцзХЮЙЪГHFDЪБМфЕФЭЦвЦЃЌШОСЯДгГІеГФЄжаЭЈЙ§ГѕЪМСмАЭЙмХХГіЃЌОМЏКЯСмАЭЙмзЊдЫЃЌЫцКѓЩјТЉжСжмЮЇЕФVATЁЃЦфжаЃЌдкHFDЮЙбј6жмЕФаЁЪѓЬхФкЃЌСмАЭв§СїгааЇЃЌСмАЭЙмЕФШОСЯЮДЗЂЩњЯджјЩјТЉЃЛдкHFDЮЙбј15жмЕФаЁЪѓЬхФкЃЌСмАЭЩјТЉЯджјдіМгЃЛдкHFDЮЙбј32жмЕФаЁЪѓЬхФкЃЌСмАЭЙмЩјТЉБфЕУИќЮЊбЯжиЃЌСмАЭвКМИКѕЭъШЋЖЈЯђЩјТЉжСVATЃЌЧвСмАЭвКТЉжївЊЗЂЩњдкСмАЭЙмбиЯпИпЖШЗжжЇКЭХХСаЮЩТвЕФЧјгђжмЮЇЃЈЭМ2aЃЌbЃЌcЃЉЃЈаЁБрзЂЃКЭМжаКьЩЋЕФЪЧОВТібЊЙмЃЌОВТібЊЙмКЭСмАЭЙмЦНааДцдкЃЉЁЃ
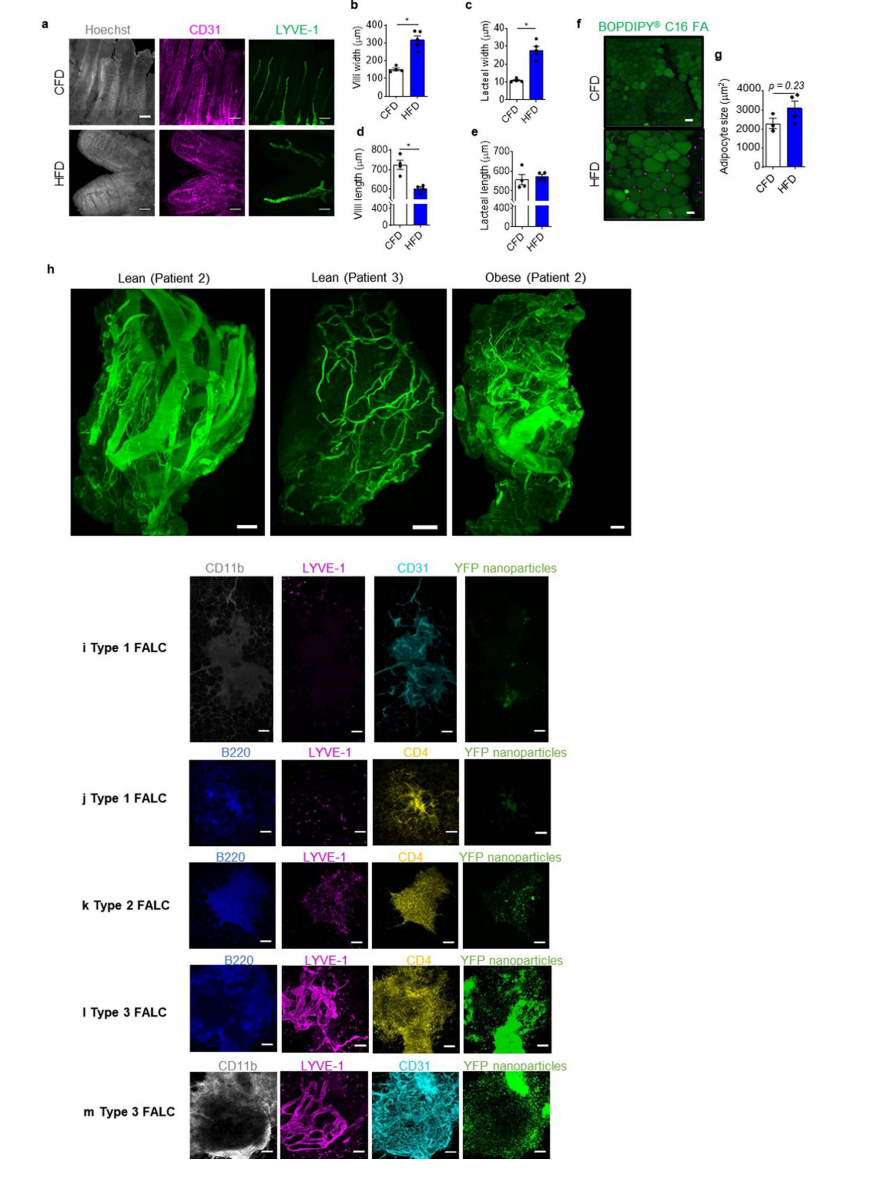
ЮЊНјвЛВНШЗЖЈГІЯЕФЄСмАЭЙІФмеЯАЪЧгЩHFDвћЪГЕМжТЕФЛЙЪЧгЩЗЪХжЫљв§Ц№ЕФЃЌбаОПШЫдБМьВтСЫCFDЮЙбјСЫ15-17жмЕФdb/dbаЁЪѓГІЯЕФЄСмАЭНсЙЙКЭЙІФмЃЈаЁБрзЂЃКdb/dbаЁЪѓЮЊleptinЪмЬхЧУГ§аЁЪѓЃЌЪГгћвжжЦЪмЫ№ЃЌНјЪГСПИпЃЌЮоашНјааHFDЮЙбјЃЌдкЦеЭЈвћЪГЯТМДПЩвђНјЪГСПИпв§ЗЂЗЪХжЃЌвђДЫПЩвдгУРДЧјЗжБОЮФЙлВьЕНЕФЯжЯѓЪЧгЩНјЪГHFDв§Ц№ЕФЃЌЛЙЪЧгЩЗЪХжв§Ц№ЕФЃЉЃЌЗЂЯжгые§ГЃCFDЮЙЪГЕФC57BL/6аЁЪѓЯрБШЃЌCFDЮЙбјЕФdb/dbаЁЪѓЬхФкВЂЮДГіЯжСмАЭЙмЗжжЇКЭСмАЭТЉвКЯжЯѓЃЛСмАЭжаЕФУтвпЯИАћМЦЪ§НсЙћвВЮоЯджјадВювьЃЈРЉеЙЪ§ОнЭМ2a-fЃЉЁЃДЫЭтЃЌdb/dbаЁЪѓЕФЦЄЯТжЌЗОзщжЏЃЈSATЃЉЬхЛ§ЯджјдіМгЃЈГЌЙ§е§ГЃHFDаЁЪѓЕФСНБЖЃЉЃЌБэУїdb/dbаЁЪѓИќШнвзЗЂЩњЭтжмжЌЗОГСЛ§ЃЈРЉеЙЪ§ОнЭМ2gЃЉЁЃживЊЕФЪЧЃЌdb/dbаЁЪѓЕФVATжиСПгыЮЙЪГHFDЕФаЁЪѓЯрЕБЃЌетаЉНсЙћБэУїГІЯЕФЄСмАЭЙмжиЙЙвРРЕгкHFDЕФЩуШыЃЌЖјгыЗЪХжЮоЙиЁЃ
ЭМ2. HFDгеЗЂЗЪХжЕФГІЯЕФЄСмАЭЙмжиЫмЕМжТСмАЭЁАЩјТЉЁБЕНГІЯЕФЄжЌЗОзщжЏ
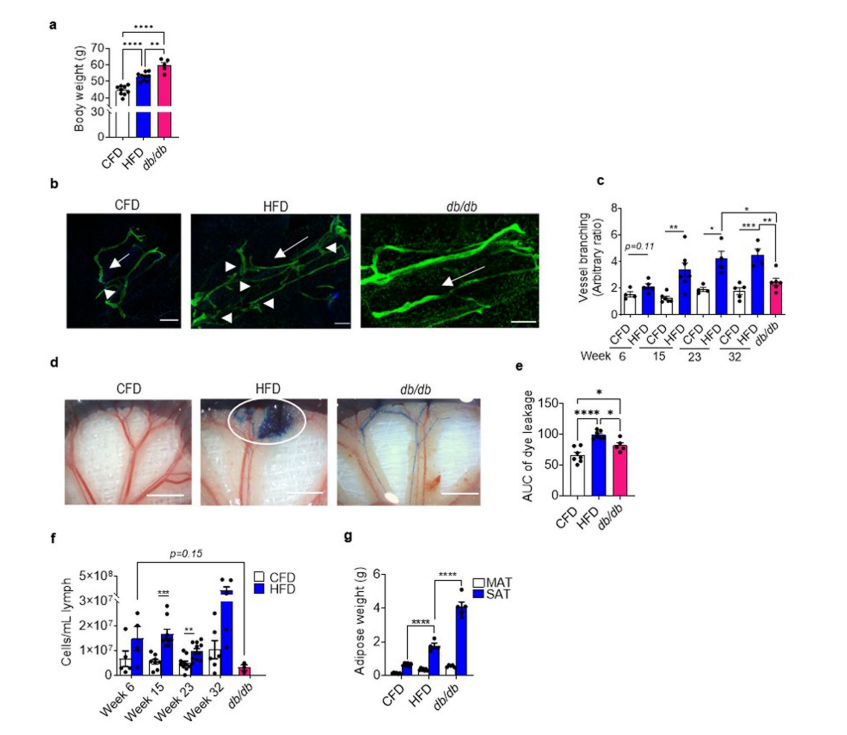
РЉеЙЪ§ОнЭМ2.Жд15-17жмСфCFDЮЙбјЕФdb/dbаЁЪѓгы6-7жмСфЮЙбјCFDЛђHFD 6ЁЂ15ЁЂ23Лђ32жмЕФC57BL/6аЁЪѓНјааГІЯЕФЄСмАЭЙІФмеЯАМьВщ
3
3.СмАЭвКЩјТЉДйНјГІЯЕФЄжЌЗОзщжЏЖбЛ§КЭвШЕКЫиЕжПЙ
МШШЛГІЯЕФЄСмАЭЙмжиЙЙвРРЕгкаЁЪѓЖдHFDЕФЩуШыЃЌЖјЗЧвРРЕгкЗЪХжЃЌФЧУДHFDгеЕМЕФГІЯЕФЄСмАЭЙмжиЙЙЪЧЗёПЩвдДйНјVATЛ§ОлВЂЕМжТвШЕКЫиЕжПЙФиЃПЮЊСЫФЃФтГІЯЕСмАЭЩјТЉЕНГІЯЕФЄжЌЗОЃЌбаОПШЫдБЪЙгУCFDКЭHFDЕФДѓЪѓЕФСмАЭвКЃЈМгШыХрбјЛљЬхЛ§ЕФ2%ЃЉЖдГЩЪьЕФ3T3-L1жЌЗОЯИАћНјааЗѕг§ЃЈаЁБрзЂЃКБОЮФЖЏЮяЪЧгУЕФДѓЪѓКЭC57аЁЪѓ, CFDКЭ HFD ЫЧЮЙаЁЪѓЫЧЮЙ6ЈC32жмЃЌДѓЪѓЫЧЮЙ6ЈC9 жмЁЃДѓЪѓЕФСмАЭвКСПЯрЖдгкаЁЪѓНЯЖрЃЌБугкЬсШЁгУгкЬхЭтЪЕбщЃЉЃЌЗЂЯжгыЦеЭЈХрбјЛљХрбјЕФ3T3-L1ЯИАћЯрБШЃЌHFDДѓЪѓСмАЭвКЗѕг§ЕФ3T3-L1ЯИАћжажЌЕЮКЭИЪгЭШ§ѕЅЖбЛ§ЯджјдіМгЃЌЖјгыCFDДѓЪѓСмАЭвКЗѕг§ЕФ3T3-L1ЯИАћЯрБШдђжЛгаЯИАћФкжЌЕЮЯджјдіМгЃЈЭМ3a-cЃЌРЉеЙЪ§ОнЭМ3a-fЃЉЃЈаЁБрзЂЃКОЁЙмCFDДІРэЕФСмАЭвКБОЩэПЩФмЛсв§Ц№жЌЗОЯИАћФкTGЕФЧсЮЂЩ§ИпЃЌЕЋЪЧдкCFDвћЪГЧщПіЯТЃЌЬхФкСмАЭВЛЛсЩјТЉЕНжмЮЇжЌЗОзщжЏжаЃЌвђДЫЯрЖдгкHFD ЖджЌЗОЕФгАЯьПЩКіТдЃЉЃЛгыЦеЭЈХрбјЛљХрбјЕФ3T3-L1ЯИАћЯрБШЃЌHFDДѓЪѓСмАЭвКЗѕг§ЕФ3T3-L1ЯИАћжЌЗОЩњГЩЯрЙиЛљвђPpargЁЂLepКЭGapdhЃЈВЛАќРЈCebpaЃЉЕФБэДяЯджјдіМгЃЈЭМ3dЃЉЃЌPnpla2ЃЈБрТыжЌНтУИATGLЕФЛљвђЃЉЕФmRNAБэДяЩ§ИпЃЈЭМ3dЃЉЃЈаЁБрзЂЃКGAPDHЃЌ3-СзЫсИЪгЭШЉЭбЧтУИЃЌЪЧВЮгыЬЧНЭНтЕФвЛжжЙиМќУИЁЃ3-СзЫсИЪгЭШЉдкGAPDHЕФДпЛЏЯТБфЮЊ1ЃЌ3-ЖўСзЫсИЪгЭЃЌетЪЧЬЧНЭНтЭООЖЮЈвЛЕФвЛВНПЩФцбѕЛЏЛЙдЗДгІЁЃИУЗДгІЪєгкЕзЮяЫЎЦНСзЫсЛЏЕФЗХФмЗДгІЃЌдкжЌЗОЫсЩњГЩЙ§ГЬжаУПдіМгвЛИіЖўЬМЕЅЮЛЖМашвЊвЛЖЈЕФФмСПЃЌЙЪGAPDHвВПЩвдгУРДзїЮЊжЌЗОЫсЩњГЩЕФЙиМќЛљвђЁЃGAPDHзїЮЊЙмМвЛљвђЃЌМИКѕдкЫљгазщжЏжаЖМИпЫЎЦНБэДяЧвБэДяСПЯрЖдКуЖЈЃЌБЛЙуЗКгУзїWestern blotЕААзжЪБъзМЛЏЕФФкВЮЁЃЕЋGAPDHдкдкЗжЛЏЙ§ГЬжаЃЌЦфБэДяСПвВЪЧБфЛЏЕФЃЌВЂЗЧвЛжБЪЪгУЃЉЁЃДЫЭтЃЌЪЙгУForskolinЃЈаЁБрзЂЃКЯймеЫсЛЗЛЏУИМЄЛюМСЃЌПЩДЬМЄжЌНтЃЉгыСмАЭвКЗжБ№ЛђЙВЭЌДЬМЄ3T3-L1ЯИАћЃЌЗЂЯжHFDДѓЪѓСмАЭвКПЩвдЯджјДйНјЛљДЁКЭForsklinДЬМЄЯТЕФжЌНтЙ§ГЬЃЛЖјCFDСмАЭвКЖдЛљДЁжЌжЪЫЎНтЫЎЦНУЛгаЯджјгАЯьЃЈЭМ3eЃЉЃЌЫЕУїHFDДѓЪѓСмАЭвКПЩвдЯджјДйНјжЌЗОжЌНтЃЈаЁБрзЂЃКЧАЮФжївЊЫЕУїСмАЭаЙТЖжСжмЮЇжЌЗОзщжЏЖјв§Ц№жЌжЪЖбЛ§ЃЛДЫДІЗЂЯжHFDСмАЭвКЛЙПЩвдДйНјжЌНтНјЖјв§Ц№вШЕКЫиЕжПЙЁЃетСНепгІИУЪЧЭЌЪБЗЂЩњЃЌЕЋЪЧДйНјЖбЛ§ЫйЖШДѓгкжЌНтЃЌЫљвдЛЙЪЧЛсв§Ц№ЖбЛ§ЃЉЁЃжЎКѓЃЌбаОПШЫдБЪЙгУСмАЭжаЕФШщУгЮЂСЃ(CM)ЁЂVLDLЃЈМЋЕЭУмЖШжЌЕААзЃЉЛђЮожЌжЪЕФЕААзЖд3T3-L1ЯИАћНјааЗѕг§ЃЌЗЂЯжДЬМЄжЌНтЕФжївЊСмАЭГЩЗжЪЧИЛКЌжЌжЪЕФШщУгЮЂСЃКЭVLDLзщЗжЃЈРЉеЙЪ§ОнЭМ3gЃЉЁЃ
гЩгкжЌЗОзщжЏжЌНтЙ§ЖШдіМгКЭЫцКѓЪЭЗХЕФгЮРыжЌЗОЫсЃЈFFAЃЉЛсдкЬхФкДйНјвШЕКЫиЕжПЙЃЌЖјвШЕКЫизїгУгкЭтжмзщжЏЪЧбЊЬЧПижЦЕФживЊОіЖЈвђЫиЃЌвђДЫбаОПШЫдБЦРЙРСЫСмАЭТЉШыVATЪЧЗёЛсЫ№КІЛњЬхЕФвШЕКЫиУєИаадЁЃдкКЌгаCFDСмАЭвКЁЂHFDСмАЭвКХрбјЛљКЭЦеЭЈХрбјЛљЕФЬѕМўЯТЃЌМьВтЛљДЁКЭвШЕКЫиДЬМЄКѓЕФ3T3-L1жЌЗОЯИАћЁЂSPF3ШЫРржЌЗОЯИАћвдМАДгЩјТЉ/ЗЧЩјТЉСмАЭЙмжмЮЇЗжРыЕФРыЬхVATЖд14C-2-ЭббѕЦЯЬбЬЧЃЈ14C-2DGЃЉЕФЩуШЁЁЃНсЙћЯдЪОЃЌгыЖдееХрбјЛљКЭCFDСмАЭвКДІРэЕФ3T3-L1жЌЗОЯИАћЯрБШЃЌЪЙгУHFDСмАЭвКДІРэКѓЃЌвШЕКЫиДЬМЄЕФ3T3-L1жЌЗОЯИАћЕФ14C-2DGЩуШЁФмСІЯджјЪмЫ№ЃЈЭМ3fЃЉЃЌИУНсЙћвВдкSPF3ШЫРржЌЗОЯИАћжаЕУЕНбщжЄЃЈРЉеЙЪ§ОнЭМ3hЃЉЁЃгыЮЙЪГCFDЕФаЁЪѓЯрБШЃЌДгЮЙЪГHFDЕФаЁЪѓЗжРыЕФVATвШЕКЫиДЬМЄКѓЕФ14C-2DGЩуШЁвВЯджјЪмЫ№ЃЛЧвгыHFDжаЗжРыЕФЗЧЩјТЉСмАЭЙмжмЮЇЗжРыЕФVATЯрБШЃЌДгЩјТЉСмАЭЙмжмЮЇЗжРыЕФVATвШЕКЫиДЬМЄКѓЕФ14C-2DGЩуШЁЪмЫ№зюбЯжиЃЈЭМ3gЃЌhЃЉЁЃБэУїHFDСмАЭвКТЉШыVATЛсДйНјОжВПГІЯЕФЄжЌЗОзщжЏЕФвШЕКЫиЕжПЙЃЌетПЩФмЕМжТШЋЩэвШЕКЫиЕжПЙЁЃ

ЭМ3. ГІЯЕФЄСмАЭТЉШыГІЯЕФЄжЌЗОзщжЏДйНјжЌЗОЯИАћЗжЛЏЁЂжЌжЪЛ§ОлКЭвШЕКЫиЕжПЙ

РЉеЙЪ§ОнЭМ3.ЮЙЪГCFDЛђHFD 6-9жмЕФSprague-DawleyДѓЪѓЕФЬхжиЁЂГІЯЕФЄСмАЭГЩЗжКЭзїгУ
4
4.COX-2КЭVEGF-CЕїНкСмАЭЙІФмКЭЦЯЬбЬЧФЭСП
ЩЯЪіНсЙћжЄУїЃЌHFDДѓЪѓЕФСмАЭвКжаКЌгаДйНјVATРлЛ§КЭвШЕКЫиЕжПЙЕФвђзгЁЃНгЯТРДЃЌбаОПШЫдБЦРЙРСЫHFDвћЪГаЁЪѓЕФСмАЭвКЪЧЗёвВАќКЌДйНјГІЯЕФЄСмАЭЙмЗжжЇКЭЩјТЉЕФвђзгЁЃЪзЯШЃЌбаОПШЫдБРћгУЯИАћЛЎКлЪЕбщжЄЪЕЃЌгыЖдееХрбјЛљЃЈМгШыCFDаЁЪѓСмАЭвКЃЉЯрБШЃЌЪЙгУ2%ЃЈvol/volЃЉЕФСмАЭвКЃЈРДздHFDЮЙбјЕФаЁЪѓЛђЗЪХжШЫШКЃЉЗѕг§КѓЃЌСмАЭЙмФкЦЄЯИАћЃЈLECЃЉЛЎКлБеКЯдіМгЃЈЭМ4a-dЃЉЃЌБэУїРДздHFDаЁЪѓКЭЗЪХжШЫШКЕФСмАЭвКФмЙЛДйСмАЭЙмЩњГЩЁЃ
дкЗЂг§КЭМВВЁЙ§ГЬжаЃЌСмАЭЙмЩњГЩЕФзюживЊНщжЪЪЧVEGF-CЁЃР§ШчЃЌдкЗЮЯйАЉЯИАћжаЙ§БэДяCOX-2ЃЌЗЂЯжCOX-2УИКЯГЩЕФЧАСаЯйЫиE2ЃЈPGE2ЃЉПЩвдЭЈЙ§еаФМВЂДЬМЄОоЪЩЯИАћЪЭЗХVEGF-CЃЌVEGF-CПЩвдгыСмАЭЙмФкЦЄЯИАћжаЕФVEGFR-3ЯрНсКЯЃЌДйНјСмАЭЙмФкЦЄЯИАћЕФдіжГЃЌНјЖјДйНјСмАЭЙмЕФЩњГЩЁЃДЫЧАЃЌгабаОПБЈЕРЃЌдкЦЄЯТжЌЗОзщжЏКЭ3T3-L1ЯИАћжаЃЌCOX-2вжжЦМСКЭVEGF-C-VEGFR3вжжЦМСЭЈЙ§ПЙбззїгУЛКНтЗЪХжКЭвШЕКЫиЕжПЙЕФЗЂЩњЁЃдкетРяЃЌбаОПШЫдБМйЩшCOX-2ЈCPGE2КЭVEGF-CЈCVEGFR3аХКХЭЈТЗЕїПиHFDЯрЙиГІЯЕФЄСмАЭЙІФмеЯАЃЌДгЖјЕМжТвШЕКЫиЕжПЙЁЃЮЊбщжЄетИіМйЩшЃЌбаОПШЫдБЭЈЙ§ЯИАћЪЕбщдкЬхЭтжЄЪЕCOX-2вжжЦМСШћРДЮєВМЁЂVEGFR3МЄУИвжжЦМСMAZ51ЃЈЭМ4bЃЉКЭЧАСаЯйЫиEP4ЪмЬхозПЙМСLY3127760ЃЈРЉеЙЪ§ОнЭМ4aЃЉФмЙЛвжжЦHFDСмАЭгеЕМЕФLECЧЈвЦЁЃжЎКѓбаОПШЫдБгжЭЈЙ§ЬхФкЪЕбщРДбщжЄетИіМйЩшЁЃбаОПШЫдБИјHFDЮЙбјаЁЪѓЗўгУCOX-2вжжЦМСШћРДЮєВМЃЈ29 mg/kg/ЬьЃЌШћРДЮєВМЛьдкHFDЫЧСЯжавЛЦ№ЫЧЮЙаЁЪѓЃЌГЦЮЊдЄЗРбаОПЃЉ15жмЃЌЗЂЯжГІЯЕФЄСмАЭжаЕФVEGF-CХЈЖШЫцзХHFDЮЙбјЖјдіМгЃЈЭМ4fЃЉЃЌШћРДЮєВМжЮСЦЕМжТСмАЭжаPGE2ЫЎЦНЯджјНЕЕЭЃЌЕЋVEGF-CЕФЫЎЦНУЛгаЯджјНЕЕЭЃЈP=0.15ЃЉЃЈЭМ4fЃЌgЃЉЃЈаЁБрзЂЃКДЫДІНЋГІЯЕФЄжЌЗОзщжЏдкКЌга2-3mg/mLНКдУИ IIЕФЯћЛЏХрбјЛљжаЧаЫщЃЌдк 37ЁуC ЯТЗѕг§35ЗжжгЁЃжЎКѓгУ100 ІЬmЯИАћЙ§ТЫЦїЙ§ТЫжЌЗОзщжЏЯћЛЏЮяЃЌЪЙгУСїЪНЯИАћвЧЗжбЁОоЪЩЯИАћ(CD45+CD11b+F4/80+)ЃЌНЋЗжбЁГіРДЕФОоЪЩЯИАћНјааELISAЗжЮіЃЌЕУЕНVEGF-CХЈЖШЃЉЁЃШћРДЮєВМжЮСЦгааЇЕиЗРжЙСЫГІЯЕФЄСмАЭЙмЗжжЇКЭЩјТЉЕФдіМгЃЈЭМ4h-kЃЉЁЃдкЮЙЪГHFDЕФаЁЪѓжаЃЌГІЯЕФЄСмАЭКЭСмАЭНсжаУтвпЯИАћЛ§ОлдіМгЃЌШћРДЮєВМжЮСЦФмЙЛвжжЦдкУтвпЯИАћдкГІЯЕФЄСмАЭвКжаЕФОлМЏЃЌЖјдкСмАЭНсжаЮозїгУЃЈРЉеЙЪ§ОнЭМ5a-fЃЉЁЃживЊЕФЪЧЃЌШћРДЮєВМЕФетаЉжЮСЦаЇЙћЫфШЛЮДИФЩЦаЁЪѓЕФПеИЙбЊЬЧЃЈp=0.06ЃЉКЭЬхжиЃЌЕЋИФЩЦСЫЛњЬхЕФЦЯЬбЬЧФЭСПЃЈOGTTЪЕбщЃЉЃЈЭМ4l-nЃЉЁЃетаЉНсЙћЫЕУїЃЌвжжЦCOX-2ЈCPGE2КЭVEGF-CЈCVEGFR3ЕФаХКХПЩФмЛсЛКНтгыHFDЯрЙиЕФГІЯЕФЄСмАЭЗжжЇдіМгЁЂСмАЭЩјТЉКЭЦЯЬбЬЧВЛФЭЪмЁЃ
ЭиеЙдФЖС
дкаЁЪѓХпЬЅжаЕФбЊЙмЯЕЭГНЈСЂКѓЃЈдМдкХпЬЅЗЂг§Й§ГЬЕФЕк10ЬьЃЉЃЌОБОВТіЧјгђПЊЪМГіЯжСмАЭЙмФкЦЄЯИАћ (LEC)ЁЃДѓЖрЪ§ LEC ЪЧгЩОВТіФкЦЄЯИАћЕФзЊЗжЛЏВњЩњЁЃLECзюГѕаЮГЩУЋЯИбЊЙмбљГѕМЖДдЃЌКѓНјвЛВНжизщаЮГЩУЋЯИбЊЙмКЭСмАЭЙмЁЃдкДЫЙ§ГЬжаЃЌгабаОПШЫдБЗЂЯжЃЌProx1 -/-аЁЪѓВЛЗЂг§ШЮКЮСмАЭбЊЙмНсЙЙЃЛдкLECжавжжЦзЊТМвђзгProx1ЕФБэДяЛсЫ№КІСмАЭЙмЕФЭъећадЃЌЕМжТСмАЭвКГЩЗжЃЈШчгЮРыжЌЗОЫсЃЉЗЂЩњаЙТЉЃЌНјЖјДйНјЧАжЌЗОЯИАћЗжЛЏЮЊжЌЗОЯИАћЃЌЫцКѓбаОПШЫдБЭЈЙ§дкLECжаЙ§БэДяProx1ЃЌЗЂЯжСмАЭЙмЭъећадЕУЕНаоИДЁЃвђДЫзЊТМвђзгPROX1вВБЛПДзїЪЧLECЕФдчЦкБъжОЮяКЭLECЕФЙиМќЕїНквђзгЁЃЩњГЄвђзг VEGFC МАЦфРвАБЫсМЄУИЪмЬх VEGFR3 ЕФаХКХДЋЕМЪЧХпЬЅЗЂЩњЙ§ГЬКЭИїжжВЁРэЬѕМўЯТLECРЉдіЕФжївЊЧ§ЖЏвђЫиЁЃVEGFCОЕААзУИЫЎНтМгЙЄКѓЃЈдкЕААзУИADAMTS3КЭЯИАћЭтЯЮНгЕААзCCBE1ЕФАяжњЯТЃЉМЄЛюVEGFR3ЃЌНјЖјДйНјСмАЭФкЦЄЯИАћЕФдіжГКЭСмАЭЙмЕФЩњГЩЁЃГ§ДЫжЎЭтЃЌVEGFC-DLL4аХКХЭЈТЗвВдкСмАЭЙмЕФЩњГЩЙ§ГЬжаЦ№зХЗЧГЃживЊЕФзїгУЃЌвжжЦVEGFCЛђVEGFR3БэДяЃЌПЩвдгааЇвжжЦШщУгЙмЃЈСмАЭЙмЕФвЛжжЃЉжаDLL4ЕФБэДяЃЌНјЖјЕМжТШщУгЙмЫ№ЩЫЃЌШщУгЙмГЄЖШЫѕЖЬЃЌЫПзДЮБзуЪ§ФПМѕЩйЃЌвђДЫГЩШЫШщУгЮЂСЃдЫЪфЕФЮЌГжвРРЕгк VEGFC-DLL4 аХКХЁЃДЫЭтЃЌгабаОПБэУїЃЌНсГІАЉЁЂЗЮАЉЁЂбЪКэАЉЁЂвШЯйАЉЁЂШщЯйАЉЕШЧжЯЎадАЉжЂЕФЗЂЩњЗЂеЙЙ§ГЬжаЃЌАЉЯИАћжаБэДяЯджјЩ§ИпЕФCOX-2ДйНјСмАЭЙмЩњГЩЁЃдкаЁЪѓШщЯйАЉФЃаЭжаЃЌАЉЯИАћжаЕФCOX-2ИпБэДяЃЌПЩвдЕМжТPGE2ЫЎЦНЩ§ИпЁЃPGE2гыEP4ЕШPGEЪмЬхНсКЯКѓЃЌЭЈЙ§ЩЯЕїVEGF-C/DРДДЬМЄСмАЭЙмФкЦЄЯИАћдіЩњЃЌНјЖјДйНјСмАЭЙмЩњГЩЛђСмАЭзЊвЦЁЃ
ВЮПМЮФЯзЃК
[1]Bernier-Latmani J, , et al. Nat Rev Gastroenterol Hepatol. 2017;14(9):510-526.
[2]Lala PK, et al. CancerMetastasis Rev. 2018;37(2-3):369-384.

ЭМ4. CoX-2ЈCPGE2КЭVEGF-CЈCVEGFR3аХКХЕїНкHFDЯрЙиЕФГІЯЕФЄСмАЭЙІФмеЯАКЭЦЯЬбЬЧВЛФЭЪм
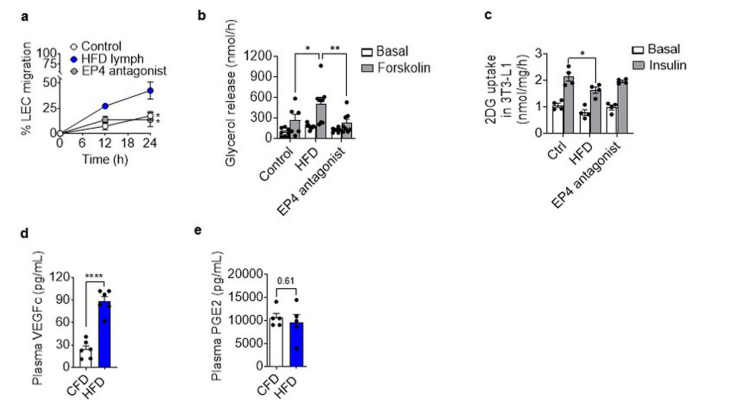
РЉеЙЪ§ОнЭМ4. EP4вжжЦЖдHFDСмАЭгеЕМЕФLECЧЈвЦЁЂжЌЗОЯИАћжЌНтКЭжЌЗОЯИАћвШЕКЫиУєИаадЕФгАЯь

РЉеЙЪ§ОнЭМ5.дЄЗРКЭжЮСЦзщГІЯЕФЄСмАЭвККЭСмАЭНсжаЕФУтвпЯИАћЛ§Ол
5
5. АаЯђвжжЦСмАЭCOX-2ЭЈЙ§МѕЩйОжВПVEGF-CЪЭЗХПЩФцзЊСмАЭЙІФмеЯА
ЛљгквдЩЯЪЕбщНсЙћЃЌПЦбаШЫдБВТВтЭЈЙ§АаЯђЛжИДГІЯЕФЄСмАЭЙІФмПЩвдФцзЊФкдрЗЪХжКЭвШЕКЫиЕжПЙЁЃЮЊСЫбщжЄетвЛМйЩшЃЌбаОПШЫдБИјаЁЪѓЮЙЪГHFD 15жмвдгеЕМжаЖШСмАЭЗжжЇЁЂСмАЭЩјТЉЁЂЗЪХжКЭвШЕКЫиЕжПЙЃЌШЛКѓдкHFDЫЧСЯжаЛьКЯШћРДЮєВМЛђАаЯђСмАЭЕФШћРДЮєВМЧАвЉЃЈCele-ProЃЉжЮСЦ7-8жмЃЈГЦЮЊжЮСЦбаОПЃЌЭМ5aЃЉЃЌвдМьВтЪЧЗёПЩвдЭЈЙ§ЪЙгУCele-proАаЯђНјШыСмАЭЕФЙІФмРДЗХДѓШћРДЮєВМЕФвЉаЇЃЌНјЖјдкНЯЕЭМССПЕФИјвЉХЈЖШЯТЛёЕУРрЫЦЛђИќКУЕФвЉЮязїгУаЇЙћЃЌдкгыШћРДЮєВМЗЂЛгЯрЭЌвЉаЇЯТПЩМѕЩйгУвЉСПЁЃЕБгУЭЌЕШИјвЉМССПЪБЃЌCele-ProжЮСЦШћРДЮєВМНјШыГІЯЕФЄСмАЭЕФзмСПдіМгСЫ10БЖвдЩЯЃЈЭМ5cЃЉЁЃдкжЮСЦбаОПжаЃЌвдCele-ProаЮЪНИјгшаЁЪѓЕФШћРДЮєВМЧАЬхМССПБШШћРДЮєВМЕЭ3.5БЖЃЌШћРДЮєВМЕФМССПгыжЎЧАЕФдЄЗРбаОПжаИјгшЕФМССПЯрЭЌ(29 mg/kg/d)(ЭМ4e-n)ЁЃЯрНЯгкЪЙгУШћРДЮєВМжЮСЦКѓбЊНЌжаИпХЈЖШЕФШћРДЮєВМЫЎЦНЃЌCele-proдкИјвЉКѓВЛНібЊНЌжаШћРДЮєВМХЈЖШКмЕЭЃЌЖјЧвдк5hКѓОЭЛљБОУЛгаЃЈРЉеЙЪ§ОнЭМ6ЃЉЃЌвђДЫЭЦВтCele-ProЕФжЮСЦаЇЙћПЩФмЪЧЙщвђгкСмАЭЬивьадаЇгІЁЃ
гыдЄЗРбаОПВЛЭЌЃЌШћРДЮєВМдкжЮСЦбаОПжаУЛгаФцзЊГІЯЕФЄСмАЭЗжжЇКЭСмАЭЩјТЉЃЌетПЩФмгыМВВЁНјеЙКѓВХПЊЪМжЮСЦгаЙиЁЃЯрБШжЎЯТЃЌЕЭМССПЕФАаЯђСмАЭCele-ProжЮСЦПЩгааЇФцзЊHFDгеЕМЕФГІЯЕФЄСмАЭЗжжЇКЭСмАЭЩјТЉШыVATЃЌжЮСЦКѓЦфЫЎЦНгыЮЙЪГCFDЕФаЁЪѓЯрЫЦЃЈЭМ5dЈCgЃЉЁЃОЁЙмШћРДЮєВМКЭCele-ProОљФмНЕЕЭСмАЭPGE2ЫЎЦНЃЌЕЋжЛгаCele-ProФмЭъШЋЯћГ§СмАЭVEGF-CЃЈЭМ5hЃЌiЃЉЁЃетНјвЛВНжЇГжСЫCOX-2КЭVEGF-CаХКХДЋЕМДйНјЗЪХжаЁЪѓГІЯЕФЄСмАЭЙмЗжжЇКЭСмАЭЩјТЉЕФМйЫЕЁЃДЫЭтЃЌГ§PGE2жЎЭтЕФЦфЫћЧАСаЯйЫиЃЈШчPCI2ЁЂ 6-ЭЊ-PGF1aЃЉдкСмАЭжаЕФЗсЖШМЋЕЭЃЌВЂЧвдкЫЧЮЙHFDЛђжЮСЦКѓХЈЖШУЛгаИФБфЃЈРЉеЙЪ§ОнЭМ7aЃЌbЃЉЃЌБэУїPGE2ЪЧВЮгыЕїНкCOX-2КЭVEGF-CаХКХЭЈТЗЕФжївЊЧАСаЯйЫиЁЃ
НгЯТРДЃЌбаОПШЫдБЗжЮіСЫДйНјГІЯЕФЄСмАЭЗжжЇКЭСмАЭТЉШыVATЕФVEGF-CЕФРДдДЁЃГІЯЕФЄСмАЭжаVEGF-CЕФРДдДПЩФмгаСНжжЃКвЛЪЧЭЈЙ§ГІЕРжБНгв§СїЛёЕУЃЌЖўЪЧЭЈЙ§ДгбЊвКжаЛёЕУЁЃШЛЖјЃЌЯрНЯгкHFDЮЙбјаЁЪѓСмАЭжаЕФVEGF-CЫЎЦННЯCFDаЁЪѓИпСЫМИЪЎжС100БЖЃЈЭМ5iЃЉЃЌЮЙЪГHFDЕФаЁЪѓбЊНЌжаЕФVEGF-CЫЎЦННіБШЮЙЪГCFDЕФаЁЪѓИпГіШ§БЖЃЌБэУїбЊвКжаЕФVEGF-CВЛЬЋПЩФмЭЈЙ§НјШыГІЯЕФЄДйНјСмАЭЙмЩњГЄ (РЉеЙЪ§ОнЭМ4d)ЁЃЖјМьВтVATКЭГІЕРжаЕФVEGF-CЫЎЦНЗЂЯжHFDЮЙбјЕФаЁЪѓVATжаVEGF-CХЈЖШЯджјдіМгЃЌЕЋГІЕРжаУЛгадіМгЃЈЭМ5i-kЃЉЁЃДЫЭтЃЌЪЙгУАаЯђСмАЭЕФCele-ProжЮСЦКѓЃЌVATКЭСмАЭжаЕФVEGF-CХЈЖШНЕЕЭжСгыЮЙЪГCFDаЁЪѓЯрЫЦЕФЫЎЦНЃЌБэУїЪЧVATКЭСмАЭжаЕФVEGF-CДйНјГІЯЕФЄСмАЭЙІФмеЯАЃЌВЂЧвCele-ProФмЙЛЭЈЙ§НЕЕЭОжВПVEGF-CЫЎЦНРДЛжИДСмАЭЙІФмЁЃдкHFDаЁЪѓЕФVATКЭГІЕРФкЃЌОоЪЩЯИАћжаЕФVEGF-CХЈЖШЯджјИпгкСмАЭЙмФкЦЄЯИАћЃЈLECsЃЉЃЈЭМ5lЃЉЃЌБэУїОоЪЩЯИАћЪЧЮЙЪГHFDаЁЪѓVATжаЩ§ИпЕФVEGF-CЕФжївЊРДдДЁЃПЦбаШЫдБНјвЛВНбаОПСмАЭЙІФмЕФЛжИДЪЧЗёЮЊОжВПОоЪЩЯИАћМѕЩйЕФНсЙћЃЌбаОПНсЙћБэУїШћРДЮєВМКЭCele-ProОљЮДНЕЕЭHFDгеЕМЕФVATжаЕФОоЪЩЯИАћЃЈCD45+ F4/80+ЃЉЕФдіМгЃЈЭМ5mЃЉЃЌетБэУїCele-ProМѕЩйСмАЭЗжжЇКЭТЉГіЪЧЭЈЙ§МѕЩйVATжаОжВПVEGF-CЪЭЗХЖјЗЧЭЈЙ§МѕЩйVATжаЕФОоЪЩЯИАћЪ§ФПЁЃ
ЭиеЙдФЖС
ШћРДЮєВМПЩЬивьадЕивжжЦЛЗбѕЛЏУИ-2ЃЈCOX-2ЃЉЁЃбзжЂДЬМЄЯТПЩгеЕМCOX-2ЩњГЩЃЌНјЖјЕМжТбзадЧАСаЯйЫиРрЮяжЪЕФКЯГЩКЭЛ§ОлЃЌгШЦфЪЧЧАСаЯйЫиE2ЃЈPGE2ЃЉЃЌв§Ц№бзжЂЁЂЫЎжзКЭЬлЭДЁЃШћРДЮєВМПЩЭЈЙ§вжжЦCOX-2зшжЙбзадЧАСаЯйЫиРрЮяжЪЕФВњЩњЃЌДяЕНПЙбзЁЂеђЭДМАЭЫШШзїгУЁЃШћРДЮєВМЧАвЉЭЈЙ§гЩШћРДЮєВМСЌНгЕНИЪгЭѕЅжїСДЕФsn-2ЮЛжУЃЈЭМ5bЃЉЁЃдкГІЕРжаЃЌДѓВПЗжЕФИЪгЭШ§жЌЭЈЙ§гыЯИАћжаЩњГЩЕФдижЌЕААзНсКЯГЩШщУгЮЂСЃЃЌНјШыСмАЭЁЃЯШЧАбаОПБэУїЃЌгыCele-proНсЙЙЯрЫЦЕФвЉЮяФмЙЛзїЮЊИЪгЭШ§ѕЅРрЫЦЮяЃЌгыГІЕРЕФИЪгЭШ§ѕЅвЛЦ№ЭЈЙ§ГІЕРНјШыСмАЭЃЌдіЧПвЉЮяЯђГІЯЕФЄСмАЭЕФзЊдЫЃЌЦфЛњжЦЪЧвЉЮяЬивьЕиДгГІЕРЭЈЙ§СмАЭЯЕЭГЖјВЛЪЧУХОВТізЊдЫЃЌШЦЙ§СЫИЮдрЪзЙ§аЇгІЃЌДгЖјЯджјЬсИпСЫПкЗўвЉЮяЕФЩњЮяРћгУЖШЁЃ
ВЮПМЮФЯзЃК
[1]Hu L, etal. Angew Chem Int Ed Engl. 2016;55(44):13700-13705.
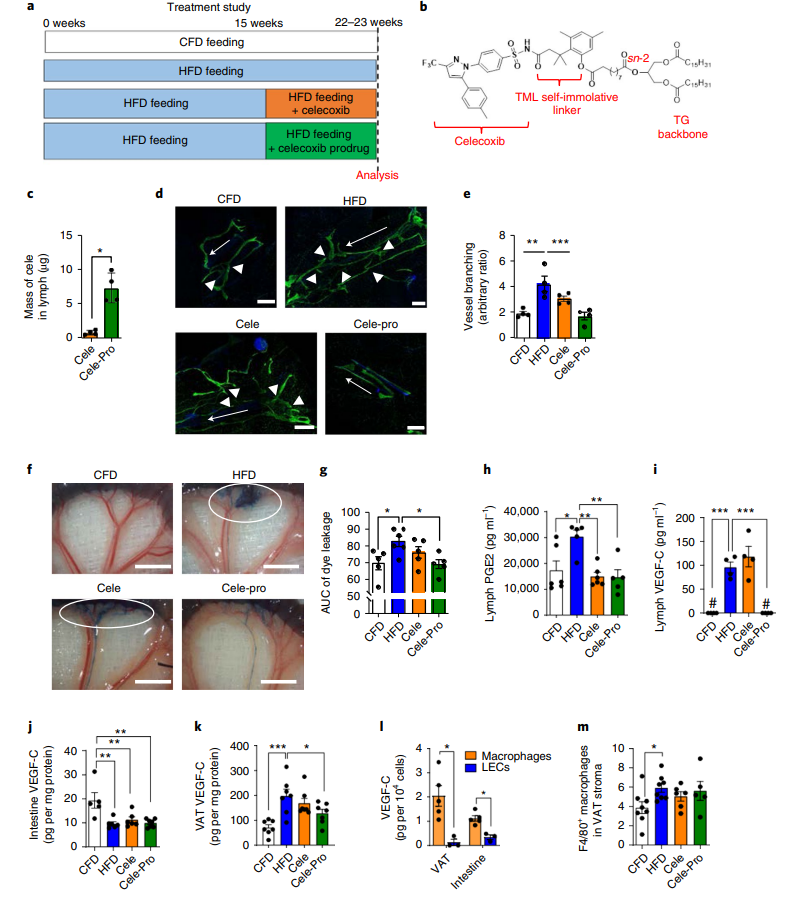 ЭМ5. СмАЭАаЯђCoX-2вжжЦЭЈЙ§МѕЩйОжВПVEGF-CЪЭЗХФцзЊHFDгеЕМЕФГІЯЕФЄСмАЭЗжжЇКЭТЉГі
ЭМ5. СмАЭАаЯђCoX-2вжжЦЭЈЙ§МѕЩйОжВПVEGF-CЪЭЗХФцзЊHFDгеЕМЕФГІЯЕФЄСмАЭЗжжЇКЭТЉГі

РЉеЙЪ§ОнЭМ6.ШћРДЮєВМЛђШћРДЮєВМЧАЬхвЉЮяЃЈCele-ProЃЉИјвЉЕФаЁЪѓЬхФкШћРДЮєВМЕФШЋЩэБЉТЖЧщПі
6
6.ГІЯЕФЄСмАЭЙІФмЕФЛжИДПЩФцзЊвШЕКЫиЕжПЙ
НгЯТРДЃЌбаОПШЫдБНјвЛВНЬНЫїСЫАаЯђвжжЦСмАЭЕФCOX-2ЪЧЗёЛсИФБфГІЯЕФЄСмАЭЛђСмАЭНсжаЕїНкбзжЂЁЂДњаЛКЭвШЕКЫиЕжПЙЕФУтвпЯИАћКЭжЌжЪДњаЛВњЮяЁЃбаОПНсЙћЯдЪОШћРДЮєВМКЭCele-ProжЮСЦОљМѕЩйГІЯЕФЄСмАЭжаЕФУтвпЯИАћЪ§СПЃЌЕЋжЛгаCele-ProФмНЋГІЯЕФЄСмАЭНсжаЕФУтвпЯИАћЪ§СПМѕЩйЕНгыCFDаЁЪѓЯрЫЦЕФЫЎЦНЃЈЭМ6aЁЂbКЭРЉеЙЪ§ОнЭМ5gЁЂhЃЉЁЃCele-ProжЮСЦЯджјдіМгГІЯЕФЄСмАЭЕФзмFFAВЂНЕЕЭЕЈЙЬДМЫЎЦН(РЉеЙЪ§ОнЭМ7c,d)ЃЌЖјСмАЭжаTG(ЭМ6c)ЁЂСзжЌКЭЦЯЬбЬЧЕФХЈЖШ(РЉеЙЪ§ОнЭМ7e,f)дкВЛЭЌзщМфУЛгаЯджјВювьЃЌетБэУїTGдкСмАЭжаЕФзЊдЫЮоУїЯдБфЛЏЃЌШщУгЙмЙІФмЕУЕНБЃСєЃЈаЁБрзЂЃКШщУгЙмЪЧПЩвдЮќЪежЌЗОЕФСмАЭЙмЁЃдкГІЕРжаЃЌжЌЗОЗжНтГЩЕЅѕЃИЪгЭКЭжЌЗОЫсЃЌБЛЮќЪеЕНаЁГІЯИАћжаЃЌжиаТКЯГЩЮЊжЌЗОЃЌВЂаЮГЩШщУгПХСЃХХГіГІЕРЁЃХХГіГІЕРЕФШщУгПХСЃПЩвдБЛШщУгЙмЮќЪеЃЌвђДЫШщУгЙмжаЕФСмАЭвКГЪЛызЧЕФШщАзЩЋЃЈИЛКЌШщУгПХСЃЕФЃЉЁЃСэЭтЃЌДЫЧАгаЮФеТБЈЕРЃЌгыCele-proНсЙЙЯрЫЦЕФвЉЮяПЩвдзїЮЊTGРрЫЦЮяЃЌгыГІЕРжаЕФTGвЛЦ№НјШыСмАЭЃЌвђДЫБОЮФМьВтСЫСмАЭжаЕФFFAКЭTGЕФКЌСПЁЃНсЙћЯдЪОЃЌCele-ProВЛгАЯьTGдкСмАЭжаЕФзЊдЫЃЉЁЃШћРДЮєВМКЭCele-ProОљНЕЕЭHFDЮЙбјаЁЪѓСмАЭЕФPGE2ЫЎЦНЃЌдкCele-ProжЮСЦКѓ(ЭМ6dКЭРЉеЙЪ§ОнЭМ7g,h)ЃЌГІЯЕФЄСмАЭжЌжЪзщбЇЗжЮіЯдЪОДњаЛЮяДгHFDЬиадЯђCFDЬиадБфЛЏЃЌЖјЪЙгУШћРДЮєВМжЮСЦКЭдЄЗРжЮСЦКѓЃЌHFDвћЪГаЁЪѓГІЯЕФЄСмАЭжЌжЪЦзУЛгаБфЛЏЃЈЭМ6dЃЌeЃЉЁЃзмЕФРДЫЕЃЌHFDвћЪГаЁЪѓСмАЭжавЛЯЕСагыЗЪХжКЭвШЕКЫиЕжПЙЯрЙиЕФЧЪжЌ(АќРЈЩёОѕЃАЗ)ЁЂчоДМКЭСзжЌ(РЉеЙЪ§ОнЭМ7g,h)гыCFDвћЪГаЁЪѓСмАЭЯрБШЯджјдіМгЁЃвђДЫЃЌетаЉжЌжЪДгСмАЭЩјТЉЕНжЌЗОзщжЏПЩФмЪЧОжВПДњаЛБфЛЏЕФвЛИіЧ§ЖЏвђЫиЁЃСэвЛИівђЫиПЩФмЪЧСмАЭжаPGE2ЫЎЦНдіМгЃЈЭМ5hЃЉЃЌШЛЖјЃЌEP4ЃЈPEG2ЕФЪмЬхЃЉозПЙМСВЂУЛгаЯджјИФЩЦHFDСмАЭДІРэЕФжЌЗОЯИАћЖд14C-2DGЕФЩуШЁЃЈРЉеЙЪ§ОнЭМ4bЃЌcЃЉЁЃ
Cele-ProжЮСЦКѓЛжИДСЫHFDЮЙбјаЁЪѓЕФVATСмАЭЙІФмЃЌЪЙДйСмАЭЙмЩњГЩЕФVEGF-CЫЎЦНе§ГЃЛЏЃЌМѕЩйСЫHFDгеЕМЕФУтвпЯИАћЛ§ОлЃЌВЂНщЕМСмАЭжЌжЪДњаЛВњЮяжиБрГЬЁЃАаЯђСмАЭЕФCele-ProИФЩЦСЫПкЗўЦЯЬбЬЧМЄКѓЕФШЋЩэбЊЬЧПижЦФмСІЃЌНЕЕЭПеИЙвШЕКЫиЫЎЦНКЭЦЯЬбЬЧДЬМЄКѓЕФвШЕКЫиЫЎЦНжСCFDЮЙбјЕФЫЎЦНЃЈЭМ6fЈCiЃЉЁЃЯрБШжЎЯТЃЌШћРДЮєВМИФЩЦСЫПкЗўЦЯЬбЬЧФЭЪмадЃЌЕЋВЂЮДЯджјИФЩЦHFDгеЕМЕФИпвШЕКЫибЊжЂЃЈЭМ6f-iЃЉЁЃбаОПШЫдБдкHFDЮЙбјЕФЕк15жмжСЕк23жмЃЌИјгшаЁЪѓВЛЭЌМССПЕФШћРДЮєВМКЭCele-ProКѓЃЌМьВщПкЗўЦЯЬбЬЧФЭСПКЭПеИЙЫЎЦНЗЂЯжЃЌCele-ProПЩЪЙHFDЮЙбјЕФаЁЪѓЛжИДжСе§ГЃбЊЬЧКЭе§ГЃбЊвШЕКЫиЫЎЦНЃЈЭМ6fЈCiЃЉЃЌЧвМССПДѓдМБШШћРДЮєВМЕЭ10БЖЃЈРЉеЙЪ§ОнЭМ8ЃЉЃЌетвВжЇГжСЫCele-ProОжВПЕїНкГІЯЕФЄСмАЭЙІФмПЩИФЩЦЯЕЭГбЊЬЧПижЦЕФМйЩшЁЃ
ОЁЙмШћРДЮєВМКЭCele-ProОљФмМѕЩйHFDв§Ц№ЕФЬхжиКЭVATжиСПдіМгЃЌЖјВЛИФБфзмЬхжиЛђжЌЗО/ЪнШтжЪСПЃЈЭМ6j-nЃЉЃЌЕЋжЛгаCele-ProПЩНЋHFDЮЙбјаЁЪѓЕФГІЯЕФЄжЌЗОзщжЏжЪСПНЕЕЭЕНгыCFDЮЙбјаЁЪѓЯрЫЦЕФЫЎЦНЃЈЭМ6lЃЉЁЃгыШћРДЮєВМШЋЩэадвжжЦCOX-2ЯрБШЃЌCele-ProАаЯђвжжЦСмАЭЕФCOX-2дкСмАЭЙІФмЁЂЬхжиЁЂФкдрЗЪХжКЭбЊЬЧПижЦЗНУцОпгазмЬхЩЯИќгХдНЕФаЇЙћЁЃЮЊНјвЛВНШЗЖЈCele-ProИФЩЦбЊЬЧПижЦЪЧНівРППГІЯЕФЄСмАЭЙІФмЛжИДЕФНсЙћЃЌЖјЗЧЖдШЋЩэДњаЛЕФгАЯьЃЌбаОПШЫдБЪЙгУШћРДЮєВМКЭCele-ProжЮСЦЮЙЪГHFDЕФаЁЪѓЃЌШЛКѓМьВтСЫЪГЮяЩуШыЁЂЗрБуФмСПХХаЙЁЂФмСПЯћКФКЭКєЮќНЛЛЛТЪЕШжИБъЃЌНсЙћЯдЪОЫљгажИБъЖМЮДЗЂЩњБфЛЏЃЌБэУїCele-ProЕФжЮСЦвцДІЪЧЭЈЙ§АаЯђОжВПСмАЭЙмЖјВЛЪЧЭЈЙ§ИФБфШЋЩэДњаЛЪЕЯжЕФЃЈЭМ6o-rЃЉЁЃ

ЭМ6. СмАЭАаЯђCoX-2вжжЦЭЈЙ§ЛжИДГІЯЕФЄСмАЭЙІФмЖјВЛЪЧИФБфШЋЩэДњаЛЙІФмРДФцзЊвШЕКЫиЕжПЙ
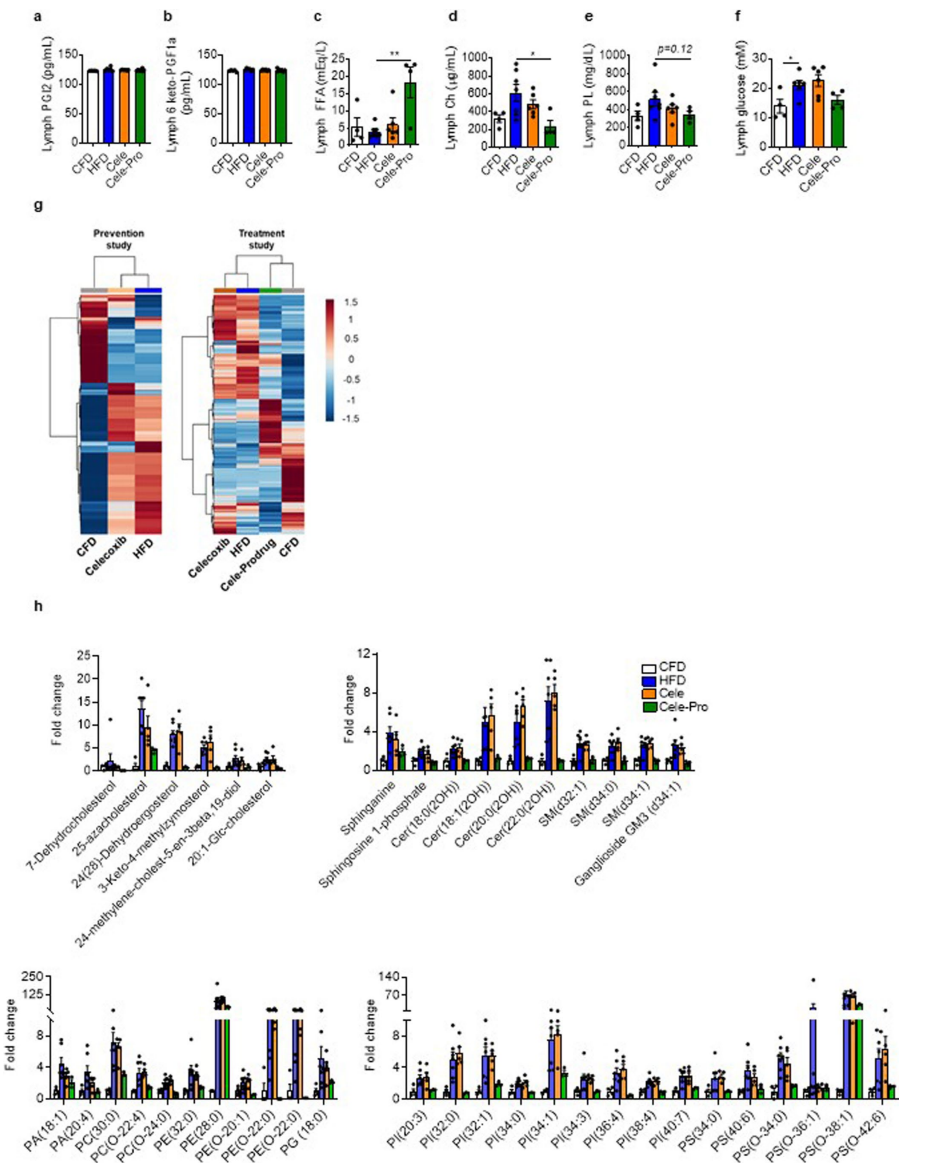
РЉеЙЪ§ОнЭМ7.ДгЮЙЪГCFDЁЂHFDЁЂHFDМгШћРДЮєВМЛђШћРДЮєВМЧАвЉЃЈCele-ProЃЉЕФаЁЪѓЛёЕУЕФГІЯЕФЄСмАЭвКжаЕФжЌжЪДњаЛВњЮя

змНс
БОЮФбаОПНсЙћБэУїГІЯЕФЄСмАЭЙІФмеЯАЪЧЗЪХжКЭвШЕКЫиЕжПЙЕФдвђКЭЧБдкжЮСЦАаБъЁЃбаОПШЫдБжЄУїСЫдкЮЙЪГHFDЕФаЁЪѓжаЃЌHFDгеЗЂЗЪХжЪБЕМжТГІЯЕФЄСмАЭЙмЗЂЩњжиЙЙЃЌГІЯЕФЄСмАЭЙмИпЖШЗжжЇЃЌСмАЭЩјТЉжСГІЯЕФЄжЌЗОзщжЏЃЌДйНјОжВПГІЯЕФЄжЌЗОзщжЏЛ§РлМАШЋЩэЕФвШЕКЫиЕжПЙЁЃГІЯЕФЄСмАЭЙІФмеЯАЪЧгЩCOX-2ЈCPGE2КЭVEGF-CЈCVEGFR3аХКХЕїПиЕФЃЌШћРДЮєВМЧАЬхвЉЮяCele-ProЭЈЙ§АаЯђвжжЦСмАЭCOX-2МѕЩйОжВПVEGF-CЪЭЗХВЂФцзЊСмАЭЙІФмеЯАЃЌзшжЙЬхжидіМгЃЌЛжИДбЊЬЧПижЦЃЌНЕЕЭЗЪХжаЁЪѓЕФИпвШЕКЫибЊжЂЁЃ
дЮФСДНгЃКhttps://www.nature.com/articles/s42255-021-00457-w
ЙизЂЮЂаХЙЋжкКХДњаЛбЇШЫ
СЫНтИќЖрДњаЛЧАбизЪбЖ

https://blog.sciencenet.cn/blog-3483272-1317285.html
ЩЯвЛЦЊЃКДњаЛбЇШЫ--Cell Metabolism ЪЎЖўдТПЏОЋбЁ
ЯТвЛЦЊЃКДњаЛбЇШЫ-- Nature Metabolism 12дТДњаЛОЋбЁ