1.
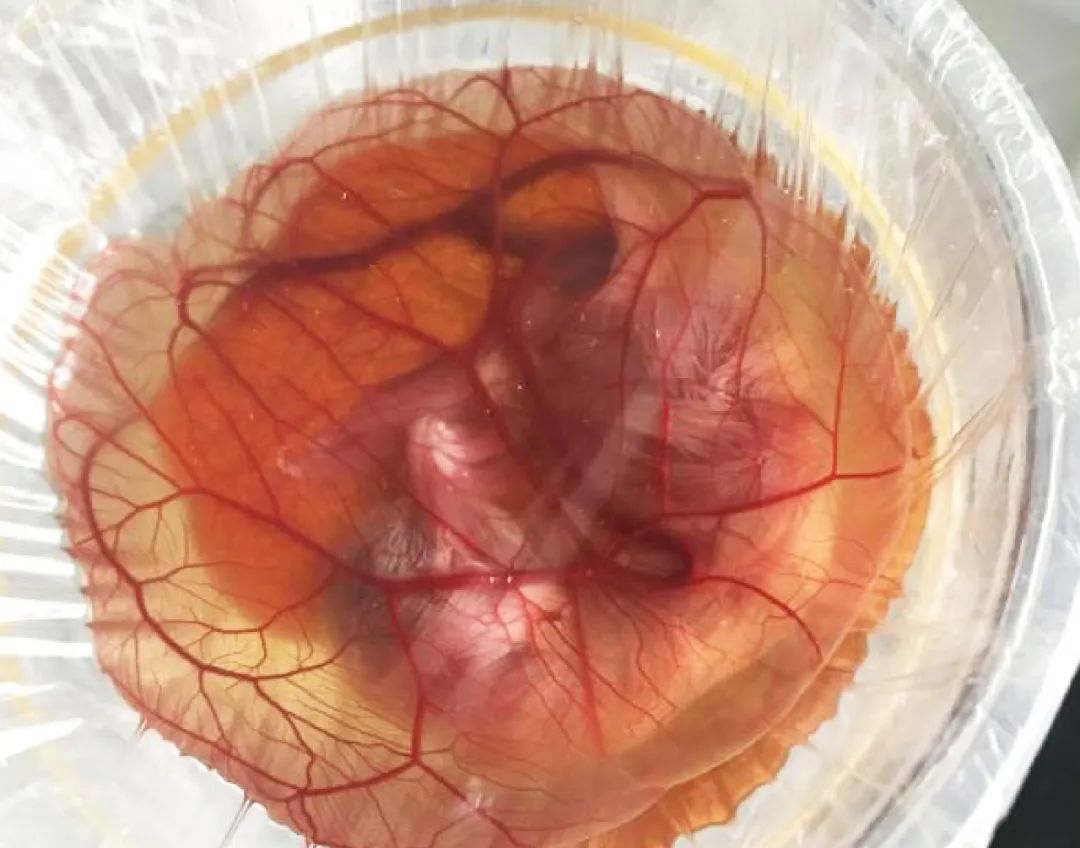
B. C. Li, et al. Relationship between PGCs Settle and Gonad Development
in the Early Chicken Embryo. Anim Biosci 2004;17(4):453-459.
2.
Ioannidis J, et al. Primary sex determination in birds depends on DMRT1
dosage, but gonadal sex does not determine adult secondary sex
characteristics. Proc Natl Acad Sci U S A. 2021 Mar
9;118(10):e2020909118.
3.
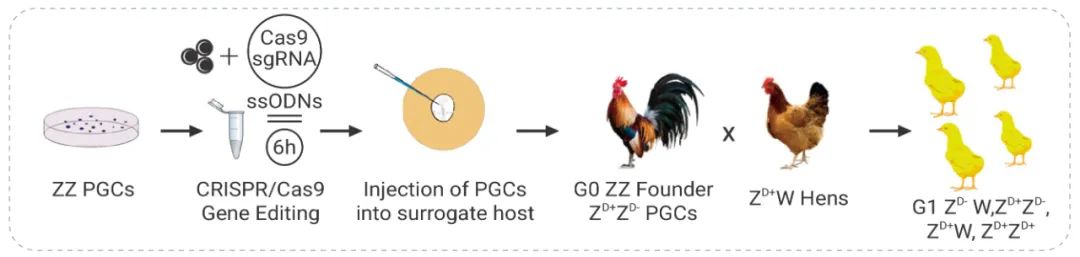
Cooper CA, et al. Generation of gene edited birds in one generation
using sperm transfection assisted gene editing (STAGE). Transgenic Res.
2017 Jun;26(3):331-347.
4. Hsu PD, et al. Development and applications of CRISPR-Cas9 for genome engineering. Cell. 2014 Jun 5;157(6):1262-1278.
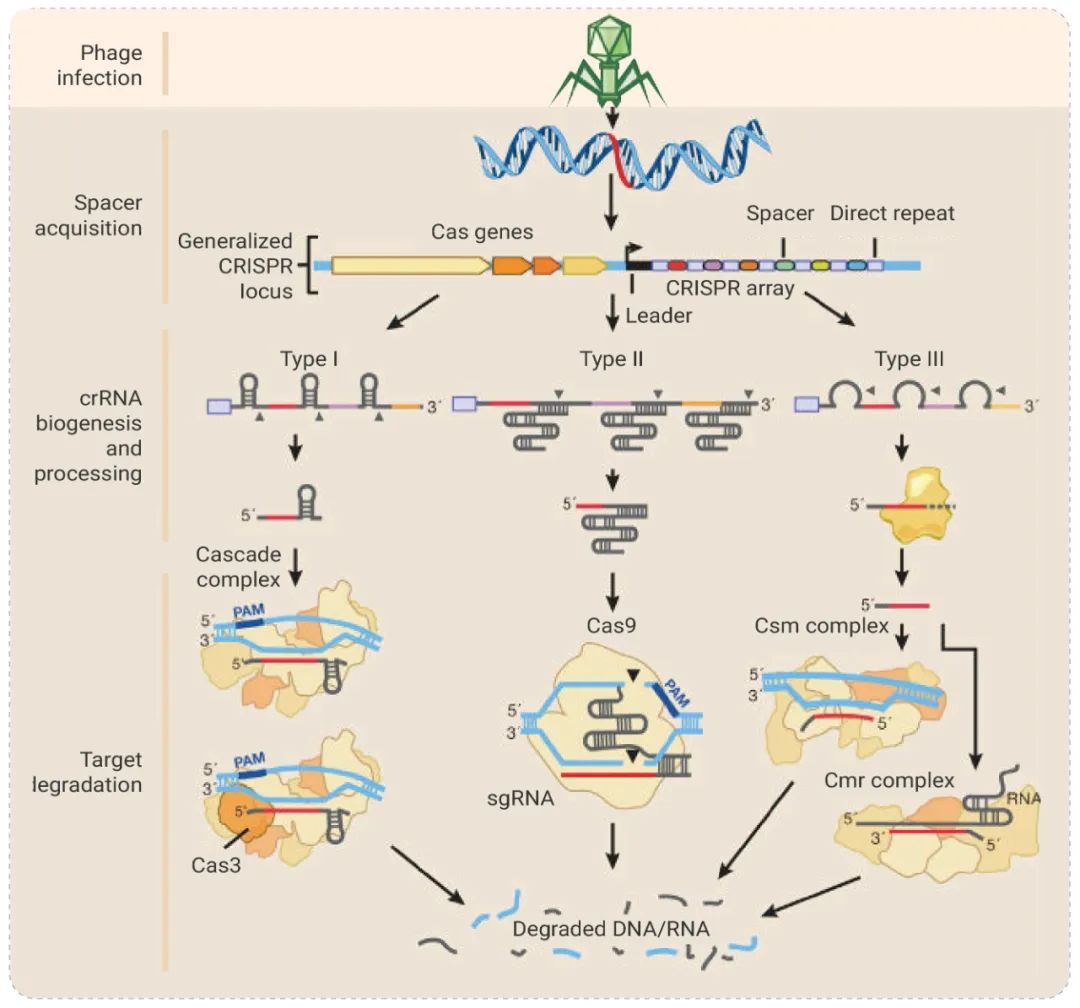
5.
Jinek M, et al. A programmable dual-RNA-guided DNA endonuclease in
adaptive bacterial immunity. Science. 2012 Aug 17;337(6096):816-21.
6.
Xu K, et al. Effective MSTN Gene Knockout by AdV-Delivered CRISPR/Cas9
in Postnatal Chick Leg Muscle. Int J Mol Sci. 2020 Apr 8;21(7):2584.
7. Long JS, et al. Species specific differences in use of ANP32 proteins by influenza A virus. Elife. 2019 Jun 4;8:e45066.
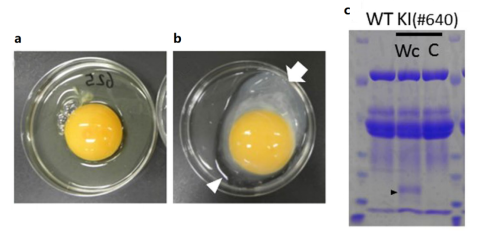
8.
Oishi I, et al. Efficient production of human interferon beta in the
white of eggs from ovalbumin gene-targeted hens. Sci Rep. 2018 Jul
5;8(1):10203.