博文
Nature综述:病毒的跨物种传播和毒力的进化(10)
|||
Nature综述:病毒的跨物种传播和毒力的进化
(10)未来的发展方向及建议
题记:
当前给全人类造成重大危害的新冠病毒可能来源于蝙蝠。新冠病毒在实现从蝙蝠到人的跨物种传播后,其毒力会如何演变?
本文介绍的原理和方法可能为我们今后进一步思考和研究相关问题提供有益的启示。
题目为《The phylogenomics of evolving virus virulence ( 病毒毒力的进化与系统基因组学 )》的综述论文于2018年12月发表在 Nature Reviews Genetics| volume 19:769-756
(本博客分10次发布该综述论文的全文,本次发布的是最后一部分:第10部分)
未来的发展方向及建议
毒力进化是进化生物学中历史最悠久的问题之一。虽然已经发展了一套坚实的理论体系,但我们仅在很少的特例中理解了推动其毒力进化的原动力,只在更少的案例中我们成功地将进化理论与个体基因组的变化联系起来。我们相信,在系统基因组框架内,毒力决定因子的实验研究和长期存在的毒力进化理论的综合,将使我们能对毒力进化产生更全面的理解。
特别是,系统基因组学方法不仅能够识别潜在的毒力决定因子,该方法还越来越多地用于新出现的病毒,而且这种分析还有助于构建阐明毒力进化的模型,相关的理论家们几十年来一直在努力构建这样的模型。
在疾病暴发期间实时基因组学的最新进展和对公共卫生干预的精确性要求的增加,可能有助于推进对病原体毒力演化的更深入的研究。我们认为,只要有相关的数据,并且在基因组学、系统发育学、流行病学和病毒毒力与适应性的实验研究之间建立起紧密的联系,就可以在系统基因组学框架内实现这一目标。因此,至关重要的是收集与病毒基因组测序同时发生的临床(即疾病症状和严重程度)和流行病学(即采样的时间和地点)的元数据(metadata),并针对一系列临床综合征(而不仅仅是针对与严重疾病相关的综合征)进行采样。
我们也强调从合适的储存宿主物种( reservoir species)收集同时发生的数据和历史数据的价值,因为这些数据可能会提供对毒力进化更完整的了解,并确定感染各种特定物种的微生物的完整类型,以及它们之间的相互作用,因为将疾病综合症归因于单个病原体常常是相当困难的。值得庆幸的是,宏基因组学(metagenomics)的进展现在让后一项任务变得可行。同样,在病毒感染中也明显缺乏良好的毒力分级方案。虽然此类方案有时可能过于简单,通常都是假设在自然界中不存在不连续的毒力类别,并纳入大量主观性指标,但粘液瘤病毒(MYXV)的案例表明,毒力分级是考虑基因型和表型之间关系的关键,这也是理解毒力进化所必需的。
最后,采用系统基因组学方法增进了对毒力进化的理解,可能有助于制定新的病原体控制和根除的策略。在疾病管理和入侵病原体的生物学控制领域,相关的方法和理论框架有可能发挥积极作用。尽管由于涉及到的病毒的巨大复杂性,预测一种新疾病何时何地出现显然是不可行的,但预测病毒在新宿主中毒力进化的总体轨迹可能是较容易达到的。
生物防治再一次提供了一个引人注目的例子。尽管存在争议,拟在澳大利亚释放鲤鱼疱疹病毒3型(cyprinid herpesvirus 3 , CyHV-3)作为对入侵的鲤鱼(Cyprinus carpio L.)的生物防治可能提供一个独特的机会,实时跟踪宿主和病毒在基因型和表型上的共同进化(co- evolution)。理论和病毒自然史都预测,CyHV-3的毒力将随时间的推移而下降,而看看这种毒力的进化如何在系统基因组数据中表现出来将是有趣和有益的。
Table 1 Examples of virulence determinants in viruses
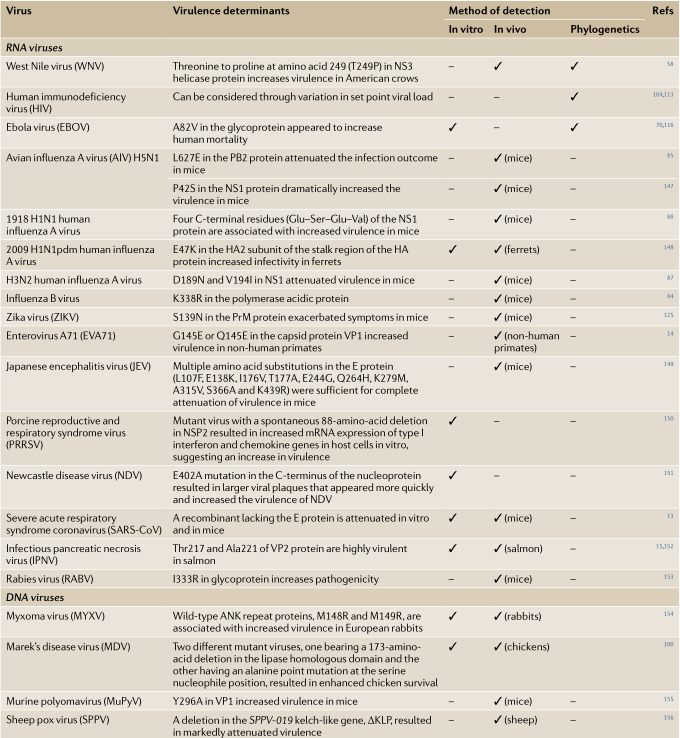
表1 病毒致病(毒力)因子举例
病毒 | 毒力因子 | 检测方法 | ||
体外 | 体内 | 系统基因组学 | ||
RNA病毒 | ||||
西尼罗河病毒(WNV) | NS3解旋酶蛋白(helicase protein)249位氨基酸由苏氨酸变脯氨酸(T249P)增加在美国乌鸦中的毒力 | ✓ | ||
人类免疫缺陷病毒(HIV) | 通过设定点病毒载量(set point viral load)的变化 | – | – | ✓ |
埃博拉病毒(EBOV) | 糖蛋白上 A82V替代似乎增加人类死亡率 | ✓ | – | ✓ |
H5N1禽流感病毒(AIV) | PB2蛋白中L627E 替代在小鼠感染中有减毒作用 NS1蛋白中的P42S替代显著增加了在小鼠中的毒力 | – | ✓ (小鼠) | – |
1918 H1N1人A型流感病毒 | NS1蛋白4 C -终端残基(Glu Ser Glu Val)增加在小鼠中的毒力 | – | (小鼠) | – |
2009 H1N1pdm人类A型流感病毒 | HA蛋白亚基柄的地区 E47K 替代增加在雪貂中的传染性 | ✓ | ✓ (雪貂) | – |
H3N2人A型流感病毒 | NS1中D189N和V194I替代导致对小鼠的毒力减弱 | – | ✓ (小鼠) | – |
B型流感病毒
| K338R替代在聚合酶中呈酸性蛋白 | – | ✓ (小鼠) | – |
Zika病毒(ZIKV) | PrM蛋白中的S139N替代加重了小鼠的症状 | – | (小鼠) | – |
肠病毒A71 (EVA71) | 衣壳蛋白VP1中的G145E或Q145E替代增加了在非人类灵长类动物中的毒力 | – | ✓ (非人类灵长类动物) | – |
日本脑炎病毒(JEV) | E蛋白中多种氨基酸的替代 | – | ✓ (小鼠) | – |
猪生殖和呼吸综合症病毒 | NSP2中自发缺失88个氨基酸的突变病毒导致了宿主细胞中I型干扰素和趋化因子基因的mRNA表达增加,表明病毒的毒力增强 | ✓ | – | – |
新城疫病毒(NDV) | ✓ | – | – | |
严重急性呼吸系统综合症冠状病毒(SARS- CoV) | 缺乏E蛋白的重组体在体外和小鼠体内是减毒的 | ✓ | ✓ (小鼠) | – |
传染性胰腺坏死病毒(IPNV) | VP2蛋白的Thr217和Ala221在鲑鱼中是高毒力的 | ✓ | ✓(鲑鱼) | – |
狂犬病毒 (RABV) | 糖蛋白中的I333R替代增加了致病性 | – | ✓ (小鼠) | – |
DNA病毒 | ||||
野生型ANK重复蛋白M148R和M149R替代,与欧洲家兔毒力增强有关 | ✓ | ✓ (兔) | ||
马立克氏病病毒(MDV) | 两种不同的突变病毒,一种在脂肪酶的同源结构域上有173个氨基酸缺失,另一种在丝氨酸亲核位点上有丙氨酸点突变,这两种病毒都提高了鸡的存活率 | ✓ | ✓ (鸡) | |
小鼠多瘤病毒(MuPyV) | VP1中的Y296A替代增强了小鼠的毒力 | – | ✓ (小鼠) | – |
羊痘病毒(SPPV) | 删除SPPV-019中的kelch样基因,ΔKLP, 导致明显的减毒 | – | ✓ (羊) | – |
这张表展示了一些突变的例子,这些突变被认为决定各种RNA和DNA病毒的毒力,以及用于鉴定这些突变的方法(在体外、体内或通过系统发育分析)。
ANK, 锚蛋白; E, 包膜; HA, 血凝素; NS1,非结构蛋白1; NSP2,非结构多蛋白2; PB2,聚合酶碱性蛋白2; pdm, 大流行; PrM, pre-membrane预膜蛋白。
相关博文:
Nature综述:病毒的跨物种传播和毒力的进化(1) 2020-03-05
Nature综述:病毒的跨物种传播和毒力的进化 (2) 2020-03-07
Nature综述:病毒的跨物种传播和毒力的进化(3) 2020-03-09
Nature综述:病毒的跨物种传播和毒力的进化(4) 2020-03-12
Nature综述:病毒的跨物种传播和毒力的进化(5) 2020-03-13
Nature综述:病毒的跨物种传播和毒力的进化 (6) 2020-03-14
Nature综述:病毒的跨物种传播和毒力的进化(7) 2020-03-15
Nature综述:病毒的跨物种传播和毒力的进化(8) 2020-03-16
Nature综述:病毒的跨物种传播和毒力的进化(9) 2020-03-17
原文全文下载:
![]() The phylogenomics of evolving virus virulence.pdf
The phylogenomics of evolving virus virulence.pdf
https://blog.sciencenet.cn/blog-347754-1224111.html
上一篇:Nature综述:病毒的跨物种传播和毒力的进化(9)
下一篇:美国将有多少人死于新冠病毒?