博文
烟草PPO家族是参与传粉的重要调控因子
||
烟草PPO家族是参与传粉的重要调控因子
多酚氧化酶(Polyphenol oxidases, PPOs)是常见的核编码含铜酶,存在于所有昆虫、真菌、脊椎动物和植物中。PPOs属于氧化还原酶家族,主要存在于叶绿体的类囊体膜中。一般来说,根据底物特异性和结构,PPOs可以可分为儿茶酚氧化酶(EC 1.10.3.1)、酪氨酸酶(EC 1.14.18.1)和漆酶(EC 1.10.3.2)。儿茶酚氧化酶催化涉及分子氧的两种不同反应:单酚的邻羟基化为邻二酚(甲酚酶活性)和邻二酚的氧化为邻醌(儿茶酚酶活性)。酪氨酸酶总是催化单酚羟基化,漆酶氧化邻二酚和对二酚形成相应的醌。
在植物组织中,PPOs可能以含有转运肽的潜伏形式存在,它们被蛋白水解裂解激活,直到它们被纳入质体。传递肽通常包含三个明显的区域:一个不带电的氨基末端区域,一个缺乏酸性残基的中心区域和一个羧基末端区域具有形成两亲性P链的潜力。在大多数已鉴定的PPOs中发现了具有保守结构域的传递肽。
PPOs是六大防御酶中的一类氧化还原酶,在防御机制中发挥重要作用,在代谢途径和生物合成中也有作用。PPOs表达的反义下调可导致植物对病原菌的超敏感性,这表明PPO介导的酚氧化在植物防御中起着关键作用。过表达PPOs的番茄植株对P . syringae增强。伤口和草食诱导的PPOs在杂交杨树中的表达支持该蛋白对害虫的防御作用。在苯丙素途径中,PPOs被认为是对香豆酸羟基化为咖啡酸的原因。在木酚油灌木中,PPOs在木酚油灌木8-8 '连接木脂素的生物合成中起着核心作用,木脂素具有有效的抗病毒、抗癌和抗氧化特性。Aureusidin合成酶(植物多酚氧化酶家族)特异性催化金色酮的氧化形成,在金鱼龙花呈现黄色中起关键作用。MicroTom番茄黄酮的生物合成与番茄多酚氧化酶(SlPPOF)有关,该酶具有类似黄酮合成酶的活性。此外,PPOs还参与甜菜素的生物合成,并参与多种生物过程,包括植物发育、细胞分化和死亡、梅勒反应、电子循环和氧调节。
已有研究表明,生殖信号通路可能与防御反应共同进化,花粉与花柱之间的相互作用与细菌防御反应有相似之处,如大多数已知的防御样蛋白已被证实主要表达于生殖器官,并在生殖过程中发挥多种作用。作为植物防御系统的成员,有报道称烟草花中有一个PPO成员(tobP1)特异表达。此外,PPOs通过调节烟草黄酮类稳态在花粉发育中发挥重要作用。这些研究表明,PPOs在植物繁殖中起着重要的作用。然而,PPOs在传粉中的潜在功能尚不清楚。
本文通过对烟叶PPO家族(研究烟叶生殖发育的重要模式系统)特征的鉴定,检测其组织表达谱和分布,探讨其在传粉中的潜在功能和机制。我们的研究将有助于加深对烟草中PPOs的科学认识,并为PPOs在植物生殖中的作用提供新的见解。
结果1:烟草PPO家族的系统发育与保守结构域分析
为了了解PPO家族的进化关系,利用NtPPO序列、来自它们祖先基因组的PPOs和来自其他植物的已发表的PPOs构建了无根系统发育树。Bootstrap值和系统发育关系将这些PPOs分为四组(I, II, III, IV), NtPPO成员分布在I组(9个NtPPOs)和II组(4个NtPPOs)(图1a)。其祖先基因组的NtPPOs与PPOs形成同源对,表明NtPPO5、NtPPO6、NtPPO8、NtPPO9、NtPPO11和NtPPO13来源于母本sylvestris, NtPPO1、NtPPO2、NtPPO3、NtPPO4、NtPPO7、NtPPO10和NtPPO12来源于父本tomentosoformis(图1a)。番茄基因组中的PPOs (PPO A/A′、PPOB、PPOC、PPOD、PPOE、PPOF)、Lycopersicon esculentum基因组(LePPO)和Solanum tuberosum基因组(StPPO)与NtPPOs (I组)归为同一组(图1a),表明它们之间的密切关系。其他PPOs (pMD-PPO2、pAPO5、VfPPO、VvPPO、PaPPO、PtPPO)和NtPPOs在不同的组中,表明它们有遥远的遗传关系(图1a)。
PPOs是核编码的含铜酶,从细胞质进入叶绿体的类囊体腔,最终定位于质体中。与典型的类囊体管腔蛋白一样,PPOs可能以潜伏形式存在,其N端含有普遍存在的转运肽。这些转运肽具有保守的结构域结构,可通过蛋白水解裂解激活。我们进一步通过多序列比对分析了NtPPOs的保守结构域,发现除了NtPPO4外,大多数NtPPO成员具有共同的结构特征(图1b)。NtPPOs的N端是一个羟基富含A.A结构域,含有许多羟基化的丝氨酸和苏氨酸残基,但酸性氨基酸很少(图1b)。高度保守的类囊体转移域(典型的类囊体腔蛋白的特征)分布在NtPPOs中,I区(“n区”)位于羟基富含A.A.A和类囊体转移域之间(图1b)。预测的假定裂解位点在Ala和Asp之间(图1b)。特别是,我们发现NtPPO4没有类囊体转移域和假定的切割位点(图1b)。在负责NtPPO蛋白催化功能的蛋白序列中分布着两个富含His的铜结合域(CuA, CuB)(图1b)。此外,所有NtPPOs共享保守结构域,包括酪氨酸酶结构域、PPO1_DWL结构域和PPO1_KFDV结构域(图1c)。
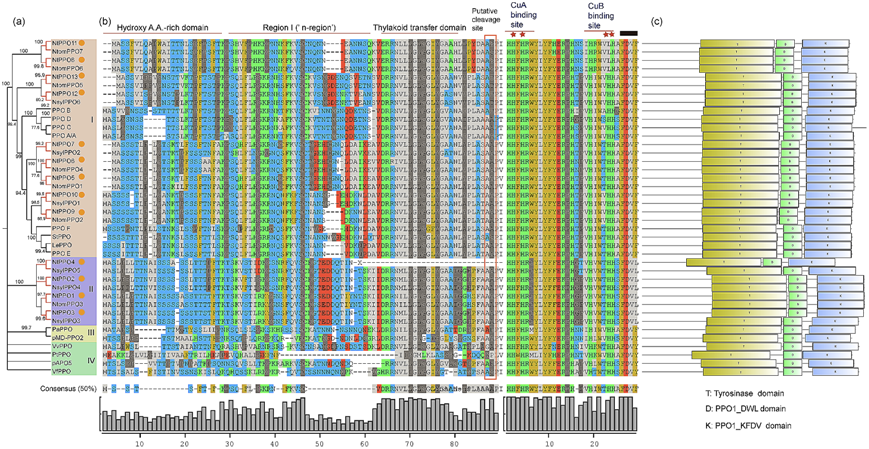
图1 烟草和其他植物的系统发育树及PPOs的保守域。(a)不同植物物种中NtPPOs和其他PPOs的系统发育树。PPO分为四组(I, II, III, IV)。NtPPO成员用橙色圆圈标记。红色分支表示一对相邻的PPOs。从其他植物物种基因组中获得的PPO序列有:富士苹果pMD-PPO2、番茄PPO A/A′、PPOB、PPOC、PPOD、PPOE、PPOF、苹果pAPO5、番茄LePPO、番茄StPPO、豆荚VfPPO、葡萄VvPPO、美洲李PaPPO、毛卡杨PtPPO。 (b)通过多序列比对推测的传递肽和功能域分析。为了实现序列间的最大同源性,引入了间隙。羟基富含A.A.A区、I区(“n区”)和类囊体转移域分别用红线标出。假定的卵裂位置用红框标出。富铜结合域CuA, CuB用红线标出。预测为铜结合位点的残基用红星标记。KFDV域用黑色方块标记。(c)保守域分析
结果2:NtPPOs启动子区顺式作用元件的多样性
为了研究NtPPO转录的潜在调控机制,我们使用PlantCARE分析了所有NtPPO成员翻译起始位点上游1.5 kb的启动子区域。预测了10个植物激素响应元件和12个非生物胁迫响应相关元件,并分析和显示了它们在NtPPOs启动子中的分布。这10个植物激素响应元件分布在大部分NtPPO成员中,包括脱落酸响应元件(ABRE)、生长素响应元件(TGAelement)、MeJA响应元件(CGTCA-motif、TGACG-motif)、乙烯响应元件(ERE)、生长素和水杨酸响应元件(as-1)和赤霉素响应元件(P-box)(图2a)。此外,在NtPPOs的启动子中还发现了水杨酸响应元件(TCAelement)、生长素响应元件(AuxRR-core)和其他赤霉素响应元件(GARE-motif)(图2a)。非生物胁迫响应相关元件,如MBS、MYB、MYC、WUN-motif、WRE3、W-box、STRE、ARE和LTR在NtPPO启动子中含量丰富(图2b)。 此外,在一些NtPPO成员的启动子中也发现了防御和应激反应元件(富含TC的重复序列)(图2b)。上述结果表明,NtPPOs的表达可能与激素信号和环境应激有关。
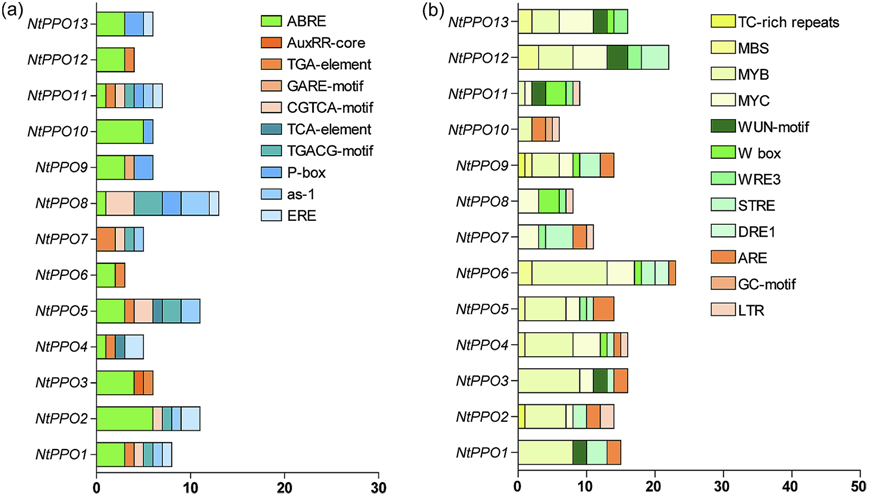
图2 NtPPO启动子中检测到的顺式元件的功能预测。(a)预测NtPPOs启动子区域的植物激素响应元件。(b)预测NtPPOs启动子区域的应力诱导和防御相关元件
结果3:NtPPOs的表达谱可能具有组织特异性
为了研究NtPPO在烟草组织中的组织特异性,我们采用RT-qPCR技术检测了NtPPO成员在烟叶根、茎、叶、雌蕊和成熟花药中的表达谱。结果表明,除NtPPO1和NtPPO3外,13个NtPPOs中有11个可以在烟叶组织中检测到(图3)。NtPPO2和NtPPO4在各组织中的表达量相对较低,而NtPPO12和NtPPO13在根中的相对表达量相对最高(图3a)。在茎叶中,NtPPO5、NtPPO6、NtPPO7、NtPPO9和NtPPO10的表达量较高(图3b, c),其中,NtPPO9在茎叶中表达量最高。此外,NtPPO9和NtPPO10在雌蕊和成熟花药中的表达水平也很高,尤其是NtPPO10(图3d和e)。值得注意的是,通过IP-LC-MS/MS分析,NtPPO9 (A0A1S3YVF2)和NtPPO10 (A0A1S4B7N5)是烟叶自花雌蕊中仅有的两个PPO成员。这些结果表明,NtPPO9和NtPPO10是最有可能在雌蕊和花粉中发挥功能的候选PPO成员,并可能在花粉管生长和授粉中发挥作用。

图3 NtPPO成员在不同组织中的表达谱。烟草根、茎、叶、雌蕊和成熟花药中NtPPOs表达的RT-qPCR检测
结果4:NtPPOs在生殖组织中大量表达
PPOs在不同组织中的表达和分布为其在组织中的功能提供了可能的线索。利用免疫荧光显微镜(IF)进一步探讨了NtPPOs在生殖组织中的表达和分布。在生长的花粉管顶端(顶端区、下顶端区(或透明区)和柄区(或剪切区)观察到强烈的绿色荧光信号(图4a)。在柱头中,在乳突细胞层中检测到微弱的绿色荧光信号(图4b)。NtPPOs在柱头和花柱的传递组织(TTS)和薄壁细胞层中表达(图4c)。在子房中,在子房壁和胚珠中观察到绿色荧光信号,荧光信号主要聚集在胚囊周围(图4d)。在雌蕊和花粉管中,NtPPOs的丰富表达和广泛分布表明其在传粉中具有潜在的功能。

图4 利用IF分析生殖组织中NtPPOs的表达和分布。阴性对照用1×PBS缓冲液代替一抗。(a)离体生长的花粉管。箭头表示花粉管顶端的NtPPOs。(b)柱头显微照片。箭头所示为乳头细胞层与传递组织界面的NtPPOs。(c)花柱显微照片。箭头显示了传递组织和实质组织中的NtPPOs。(d)卵巢显微照片。箭头所示为子房壁和胚珠中的NtPPOs。Ct,结缔组织; Aw,花药壁; Pc,乳头状细胞; Pt,实质组织; TTS,传输组织
结果5:NtPPOs表达在自相容柱头/花柱中被激活
为了进一步探讨NtPPOs是否在授粉后花粉管生长中发挥作用,我们在授粉后2.5、5、10和15h (HAP)通过IF和酶活性测定检测了NtPPOs在烟草自花(自亲和)柱头/花柱中的表达。结果表明,在2.5 HAP时,未授粉的柱头/花柱与自授粉的柱头/花柱之间的NtPPO信号(绿色荧光)强度差异不显著,且在5、10和15 HAP时,未授粉的柱头/花柱中的NtPPO信号比自授粉的柱头/花柱中的NtPPO信号弱(图5a1-a3, b和c)。在柱头中,NtPPO信号在乳头细胞层中很少观察到,主要分布在软组织组织中(图5a1)。在2.5、5和15 HAP时,NtPPO信号分布在柱头TTS中(图5a2)。有趣的是,授粉10 h后,在TTS中很少检测到NtPPO信号,而分布在TTS的边界(图5a2)。在花柱中,NtPPOs主要分布在TTS的边界,很少在TTS的中间被检测到(图5a3)。烟草自花雌蕊中NtPPOs酶活性在15 HAP时最高(图5d)。这些结果表明,授粉后NtPPOs的表达被激活,并且授粉后NtPPOs积累更多,酶活性也增加,进一步证实了NtPPOs在授粉后花粉管生长中的潜在作用。

图5 NtPPOs在烟草自花柱头/花柱中的表达及分布分析。阴性对照用1×PBS缓冲液代替一抗。(a)在2.5、5、10和15 HAP条件下,烟粉芸的柱头和花柱形态以及未授粉的柱头/花柱和自花授粉的柱头/花柱中NtPPO含量和分布的IF分析。NP,未授粉; HAP,授粉后小时数; Pc,乳头状细胞; TTS,传输组织; Pt,实质组织。红框表示观测地点。红色箭头表示NtPPO荧光信号。标尺:5 0 μm。(b-c)绿色荧光信号强度定量。误差条表示三次重复的标准差,** P < 0.01,学生t检验。(d)烟草NP柱头/花柱和自花柱头/花柱中NtPPOs的活性。进行了3次生物重复。不同字母的误差条差异有统计学意义(P < 0.05)。数据采用方差分析
结果6:NtPPOs基因敲除会影响授粉后的花粉生长和果实质量
为了研究NtPPOs在植物生殖中的功能,我们构建了针对烟草中所有NtPPOs的RNAi系。WT和两个NtPPOs沉默系(NtPPO-RNAi3/6)自花授粉5和8 h,用苯胺蓝染色,在光学显微镜下观察。结果表明,在5和8 HAP时,NtPPO-RNAi3/6系雌蕊花粉管的生长速度明显低于WT雌蕊(图6a);半离体试验还显示,NtPPO-RNAi3/6品系雌蕊花粉管生长速率较WT降低(图6d和e),果实重量明显轻于WT(图6f和g)。这些数据表明,NtPPO-RNAi3/6品系参与了授粉和果实发育。
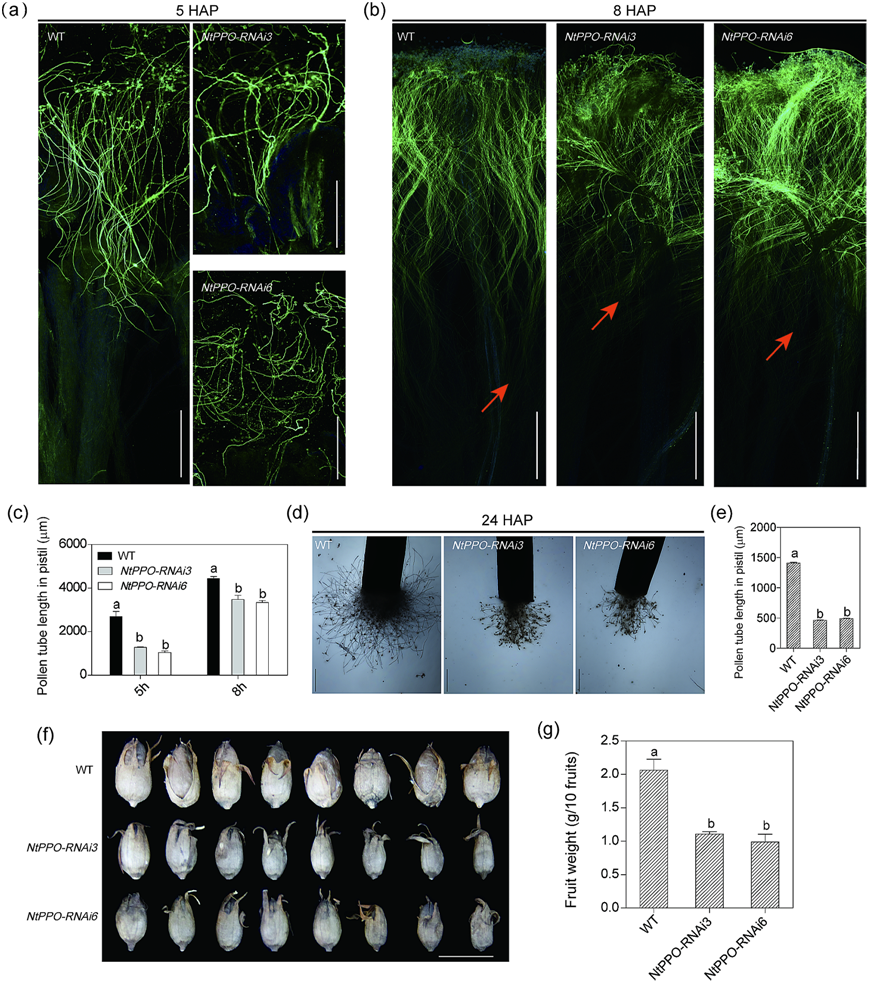
图6 NtPPOs影响雌蕊花粉生长和果实重量。(a-b)花柱第5和第8位花粉管。自花授粉后数小时。(c)自花雌蕊花粉管长度的定量。在三个生物重复中测量N = 15个授粉雌蕊,误差条表示标准差。数据采用方差分析,P < 0.05。(d-e) 24 HAP时雌蕊花粉管生长的半离体试验。在三个生物重复中测量N = 15个授粉雌蕊,误差条表示标准差。数据采用方差分析,P < 0.05。(f-g)成熟果实和果实重量量化。在三个生物重复中测量N = 30个果实; 每组含有10个水果。误差条表示三个生物重复的标准差。数据采用方差分析,P < 0.05
结果7:敲除NtPPOs后,嘌呤和黄酮类化合物在授粉雌蕊中积累
PPOs是一类酶,通过催化物质的合成和代谢发挥作用。因此,为了探究NtPPOs在传粉中的作用机制,我们采用非靶向代谢组学技术检测了NtPPO-RNAi系与WT系烟草授粉雌蕊中代谢物的差异。结果显示,NtPPO-RNAi系与WT系共有211种代谢物存在差异,其中累积和减少的分别为169和42个(图7a)。NtPPO-RNAi系与WT在嘌呤代谢、黄酮和黄酮醇生物合成、甘油磷脂代谢三个途径上存在显著差异(图7b)。差异代谢物包括dAMP、鸟苷、鸟嘌呤、腺嘌呤、3 ' 5 '-环GMP和山奈酚、黄芪甲苷、奥丁素、异槲皮素等主要富集于嘌呤代谢途径和黄酮及黄酮醇生物合成途径。在苯丙类生物合成途径中,差异代谢物磷酸胆碱和磷酸酰基甘油胆碱减少(图7c)。综上所述,这些结果表明NtPPOs主要通过嘌呤和类黄酮化合物在授粉中起作用。
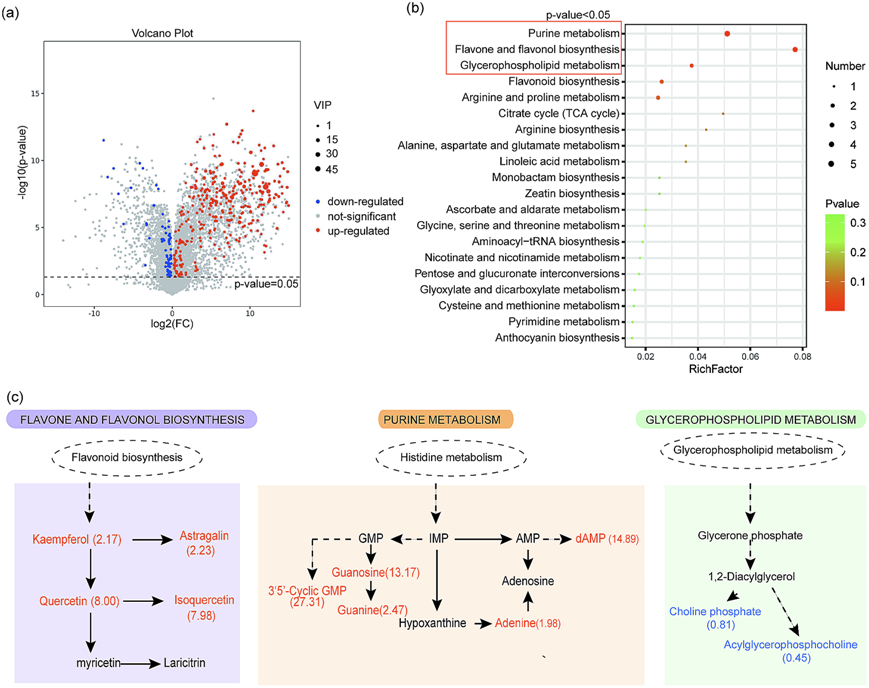
图7 敲低NtPPOs后授粉雌蕊差异代谢物分析。(a)火山图显示了NtPPO-RNAi系与WT在授粉雌蕊中差异代谢物。(b)气泡图显示了NtPPO-RNAi系与WT在授粉雌蕊中差异代谢物富集的KEGG通路,红框表示NtPPO-RNAi系与WT在KEGG通路上差异显著,P < 0.05。(c)差异显著途径中的差异代谢物。代谢物用颜色表示NtPPO-RNAi细胞系中相对于WT的丰度变化: 红色表示丰度增加和上调; 蓝色表示减少和下调; 黑色表示没有变化。代谢物后的定量值代表折叠变化(NtPPO-RNAi系/野生型)。虚线箭头表示多个酶催化反应。dAMP,脱氧腺苷单磷酸; AMP, 5 ' -单磷酸腺苷; IMP,肌苷单磷酸; GMP,鸟苷单磷酸
综上所述,我们分析了NtPPOs的系统发育关系、保守结构域和顺式作用元件,并探讨了它们在烟草生殖组织中的表达、分布和定位。敲除NtPPOs后,雌蕊花粉管生长速度减慢,果实重量降低,嘌呤和黄酮类化合物在授粉雌蕊中积累。我们的实验结果表明,NtPPOs是参与传粉的重要调控因子,并将为PPOs在生殖中的作用提供新的见解。
参考文献
[1] Wei X, Tao K, Liu Z, Qin B, Su J, Luo Y, Zhao C, Liao J, Zhang J. The PPO family in Nicotiana tabacum is an important regulator to participate in pollination. BMC Plant Biol. 2024 Feb 9;24(1):102. doi: 10.1186/s12870-024-04769-3.
以往推荐如下:
5. EMT标记物数据库:EMTome
8. RNA与疾病关系数据库:RNADisease v4.0
9. RNA修饰关联的读出、擦除、写入蛋白靶标数据库:RM2Target
13. 利用药物转录组图谱探索中药药理活性成分平台:ITCM
19. 基因组、药物基因组和免疫基因组水平基因集癌症分析平台:GSCA
22. 研究资源识别门户:RRID
24. HMDD 4.0:miRNA-疾病实验验证关系数据库
25. LncRNADisease v3.0:lncRNA-疾病关系数据库更新版
26. ncRNADrug:与耐药和药物靶向相关的实验验证和预测ncRNA
28. RMBase v3.0:RNA修饰的景观、机制和功能
29. CancerProteome:破译癌症中蛋白质组景观资源
30. CROST:空间转录组综合数据库

https://blog.sciencenet.cn/blog-571917-1426790.html
上一篇:RNA的“群体控制”:生物学中普遍存在的主题
下一篇:MarkerMap:单细胞研究的非线性标志物选择